

Dislocation of HMG-CoA reductase and Insig-1, two polytopic endoplasmic reticulum proteins, en route to proteasomal degradation
- PMID: 19458199
- PMCID: PMC2710830
- DOI: 10.1091/mbc.e08-09-0953
Dislocation of HMG-CoA reductase and Insig-1, two polytopic endoplasmic reticulum proteins, en route to proteasomal degradation
Abstract
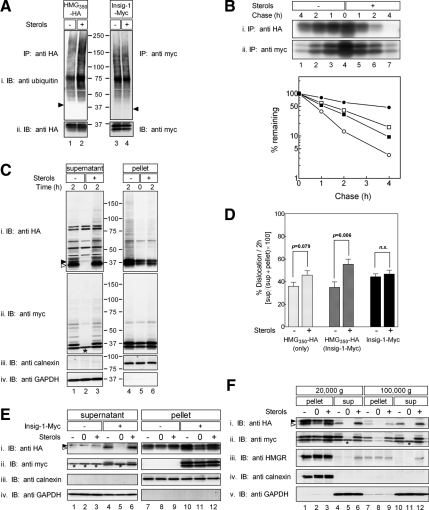
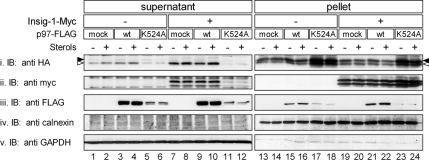
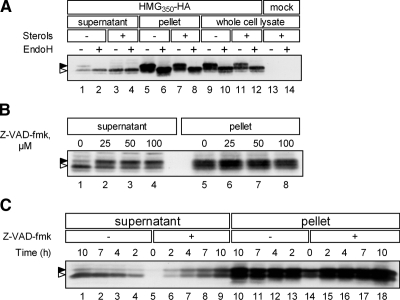
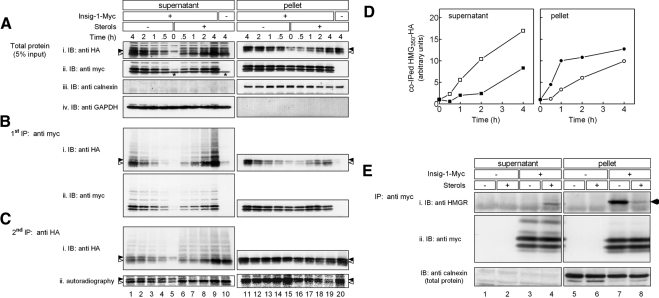
The endoplasmic reticulum (ER) glycoprotein HMG-CoA reductase (HMGR) catalyzes the rate-limiting step in sterols biosynthesis. Mammalian HMGR is ubiquitinated and degraded by the proteasome when sterols accumulate in cells, representing the best example for metabolically controlled ER-associated degradation (ERAD). This regulated degradation involves the short-lived ER protein Insig-1. Here, we investigated the dislocation of these ERAD substrates to the cytosol en route to proteasomal degradation. We show that the tagged HMGR membrane region, HMG(350)-HA, the endogenous HMGR, and Insig-1-Myc, all polytopic membrane proteins, dislocate to the cytosol as intact full-length polypeptides. Dislocation of HMG(350)-HA and Insig-1-Myc requires metabolic energy and involves the AAA-ATPase p97/VCP. Sterols stimulate HMG(350)-HA and HMGR release to the cytosol concurrent with removal of their N-glycan by cytosolic peptide:N-glycanase. Sterols neither accelerate dislocation nor stimulate deglycosylation of ubiquitination-defective HMG(350)-HA((K89 + 248R)) mutant. Dislocation of HMG(350)-HA depends on Insig-1-Myc, whose dislocation and degradation are sterol independent. Coimmunoprecipitation experiments demonstrate sterol-stimulated association between HMG(350)-HA and Insig-1-Myc. Sterols do not enhance binding to Insig-1-Myc of HMG(350)-HA mutated in its sterol-sensing domain or of HMG(350)-HA((K89 + 248R)). Wild-type HMG(350)-HA and Insig-1-Myc coimmunoprecipitate from the soluble fraction only when both proteins were coexpressed in the same cell, indicating their encounter before or during dislocation, raising the possibility that they are dislocated as a tightly bound complex.
Figures
Similar articles
-
Sequential actions of the AAA-ATPase valosin-containing protein (VCP)/p97 and the proteasome 19 S regulatory particle in sterol-accelerated, endoplasmic reticulum (ER)-associated degradation of 3-hydroxy-3-methylglutaryl-coenzyme A reductase.J Biol Chem. 2014 Jul 4;289(27):19053-66. doi: 10.1074/jbc.M114.576652. Epub 2014 May 24. J Biol Chem. 2014. PMID: 24860107 Free PMC article.
-
Sterol-induced dislocation of 3-hydroxy-3-methylglutaryl coenzyme A reductase from membranes of permeabilized cells.Mol Biol Cell. 2013 Nov;24(21):3300-8. doi: 10.1091/mbc.E13-03-0157. Epub 2013 Sep 11. Mol Biol Cell. 2013. PMID: 24025715 Free PMC article.
-
Ubiquitin is conjugated by membrane ubiquitin ligase to three sites, including the N terminus, in transmembrane region of mammalian 3-hydroxy-3-methylglutaryl coenzyme A reductase: implications for sterol-regulated enzyme degradation.J Biol Chem. 2004 Sep 10;279(37):38184-93. doi: 10.1074/jbc.M405935200. Epub 2004 Jul 7. J Biol Chem. 2004. PMID: 15247208
-
Underlying mechanisms for sterol-induced ubiquitination and ER-associated degradation of HMG CoA reductase.Semin Cell Dev Biol. 2018 Sep;81:121-128. doi: 10.1016/j.semcdb.2017.10.019. Epub 2017 Nov 7. Semin Cell Dev Biol. 2018. PMID: 29107682 Free PMC article. Review.
-
Control of cholesterol synthesis through regulated ER-associated degradation of HMG CoA reductase.Crit Rev Biochem Mol Biol. 2010 Jun;45(3):185-98. doi: 10.3109/10409238.2010.485605. Crit Rev Biochem Mol Biol. 2010. PMID: 20482385 Free PMC article. Review.
Cited by
-
Differential regulation of HMG-CoA reductase and Insig-1 by enzymes of the ubiquitin-proteasome system.Mol Biol Cell. 2012 Dec;23(23):4484-94. doi: 10.1091/mbc.E12-08-0631. Epub 2012 Oct 19. Mol Biol Cell. 2012. PMID: 23087214 Free PMC article.
-
Dual functions of Insig proteins in cholesterol homeostasis.Lipids Health Dis. 2012 Dec 18;11:173. doi: 10.1186/1476-511X-11-173. Lipids Health Dis. 2012. PMID: 23249523 Free PMC article. Review.
-
Sterol-induced dislocation of 3-hydroxy-3-methylglutaryl coenzyme A reductase from endoplasmic reticulum membranes into the cytosol through a subcellular compartment resembling lipid droplets.J Biol Chem. 2010 Jun 18;285(25):19288-98. doi: 10.1074/jbc.M110.134213. Epub 2010 Apr 20. J Biol Chem. 2010. PMID: 20406816 Free PMC article.
-
Loss of N-Glycanase 1 Alters Transcriptional and Translational Regulation in K562 Cell Lines.G3 (Bethesda). 2020 May 4;10(5):1585-1597. doi: 10.1534/g3.119.401031. G3 (Bethesda). 2020. PMID: 32265286 Free PMC article.
-
Subcellular Fractionation Analysis of the Extraction of Ubiquitinated Polytopic Membrane Substrate during ER-Associated Degradation.PLoS One. 2016 Feb 5;11(2):e0148327. doi: 10.1371/journal.pone.0148327. eCollection 2016. PLoS One. 2016. PMID: 26849222 Free PMC article.
References
-
- Baker B. M., Tortorella D. Dislocation of an ER membrane glycoprotein involves the formation of partially dislocated ubiquitinated polypeptides. J. Biol. Chem. 2007;282:26845–26856. - PubMed
-
- Bar-Nun S. The role of p97/Cdc48p in endoplasmic reticulum-associated degradation: from the immune system to yeast. Curr. Top. Microbiol. Immunol. 2005;300:95–125. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
