

Review
doi: 10.1038/nature08144.
The structure and function of G-protein-coupled receptors
Affiliations
- PMID: 19458711
- PMCID: PMC3967846
- DOI: 10.1038/nature08144
Item in Clipboard
Review
The structure and function of G-protein-coupled receptors
Nature.
.
Abstract
G-protein-coupled receptors (GPCRs) mediate most of our physiological responses to hormones, neurotransmitters and environmental stimulants, and so have great potential as therapeutic targets for a broad spectrum of diseases. They are also fascinating molecules from the perspective of membrane-protein structure and biology. Great progress has been made over the past three decades in understanding diverse GPCRs, from pharmacology to functional characterization in vivo. Recent high-resolution structural studies have provided insights into the molecular mechanisms of GPCR activation and constitutive activity.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
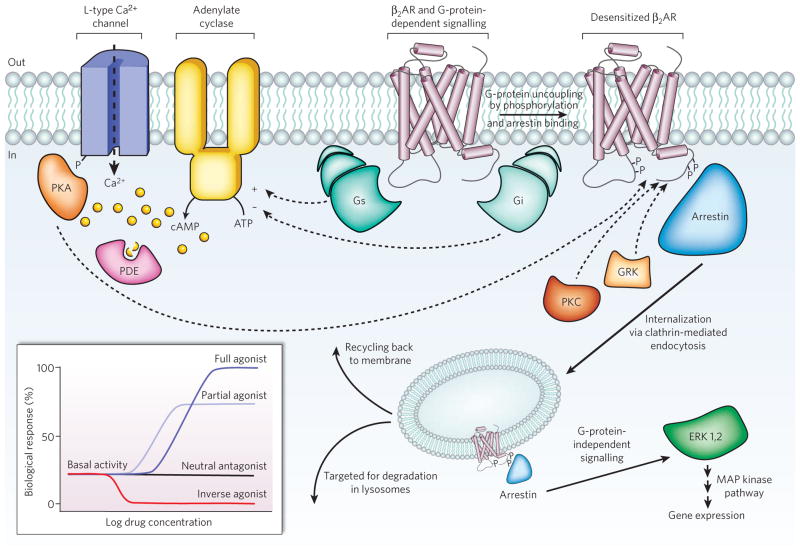
Diverse signalling pathways regulated by the type 2 beta adrenergic receptor (β2AR). The β2AR can activate two G proteins, Gαs and Gαi (part of the Gs and Gi heterotrimers, respectively), which differentially regulate adenylate cyclase. Adenylate cyclase generates cyclic AMP (cAMP), which activates protein kinase A (PKA), a kinase that regulates the activity of several cellular proteins including the L-type Ca2+ channel and the β2AR. cAMP second messenger levels are downregulated by specific phosphodiesterase proteins (PDEs). Activation of the β2AR also leads to phosphorylation by a G-protein-coupled receptor kinase (GRK) and subsequent coupling to arrestin. Arrestin is a signalling and regulatory protein that promotes the activation of extracellular signal-regulated kinases (ERK), prevents the activation of G proteins and promotes the internalization of the receptor through clathrin-coated pits. PKC, protein kinase C. The inset shows classification of ligand efficacy for GPCRs. Many GPCRs exhibit basal, agonist-independent activity. Inverse agonists inhibit this activity, and neutral antagonists have no effect. Agonists and partial agonists stimulate biological responses above the basal activity. Efficacy is not directly related to affinity; for example, a partial agonist can have a higher affinity for a GPCR than a full agonist.
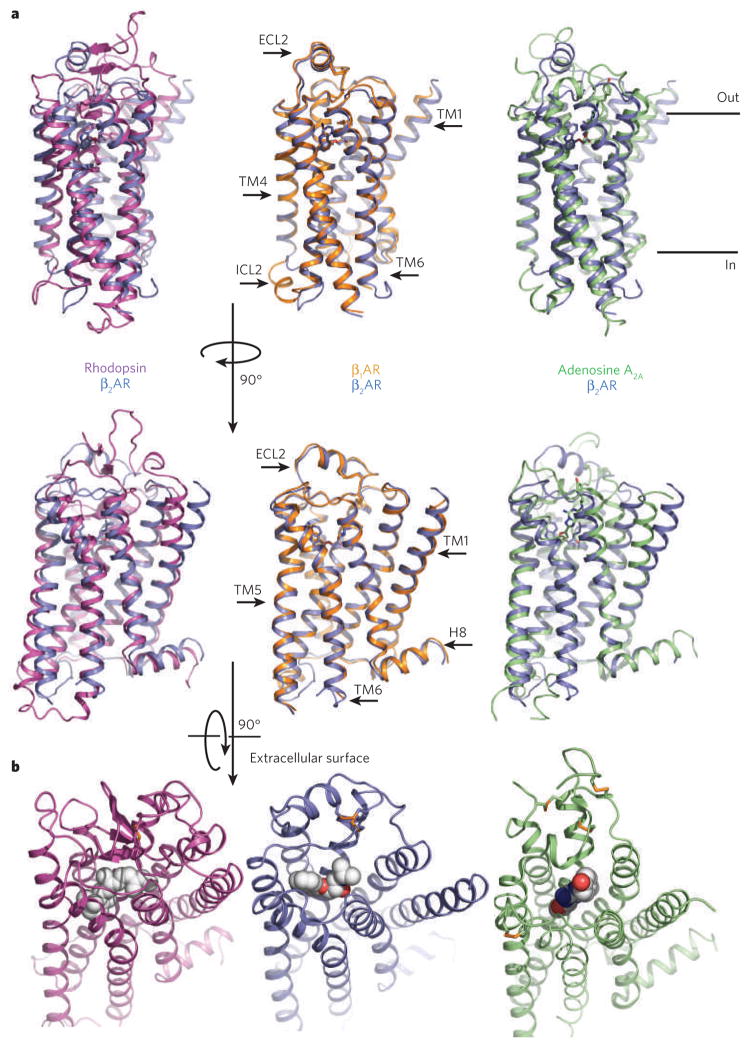
a, Bovine rhodopsin (purple), avian β1AR (orange) and human A2A adenosine receptor (green) are each superimposed on the human β2AR structure (blue). The extracellular loop 2 (ECL2), intracellular loop 2 (ICL2), cytoplasmic helix 8 (H8) and several of the transmembrane segments are indicated on one of the structures. The greatest diversity in these structures lies in the extracellular ends of the transmembrane helices and the connecting loops. b, Extracellular views of rhodopsin, the β2AR and the A2A adenosine receptor. The ligands are shown as spheres.
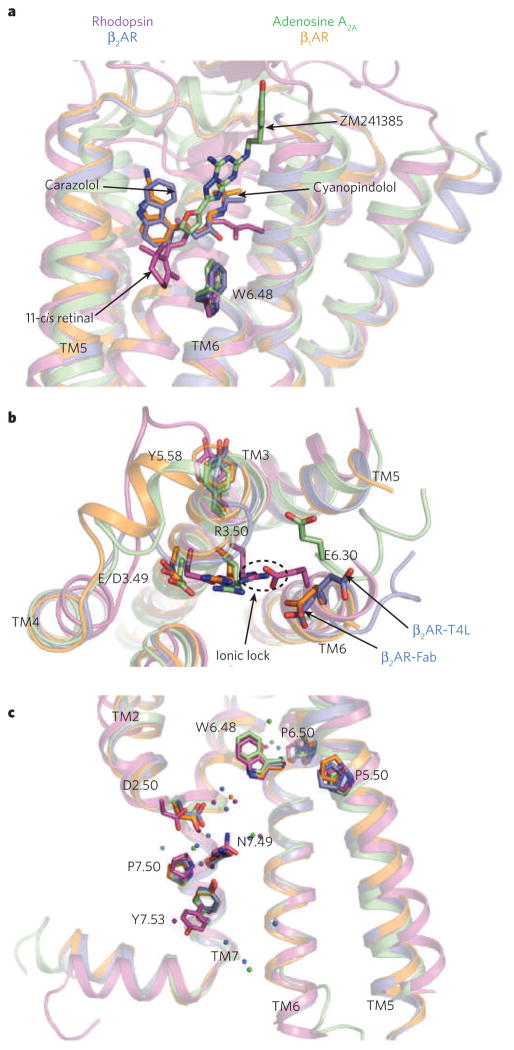
a, The locations of bound ligands for the four superimposed receptor structures bovine rhodopsin (purple, bound to 11-cis retinal), avian β1AR (orange, bound to cyanopindolol), human A2A adenosine receptor (green, bound to ZM241385) and human β2AR (blue, bound to carazolol) are shown. W6.48 is the key residue of the rotamer toggle switch. TM, transmembrane segment. b, The ionic-lock residues at the cytoplasmic end of TM3 (R3.50 and E/D3.49), and TM6 (E6.30) are shown for the same four structures. R3.50 engages Y5.58 on TM5, rather than E6.30 on TM6 in the opsin ‘active state’. The rotameric position of E6.30 differs for the two β2AR structures. c, The location of several highly conserved residues around a cluster of water molecules (coloured spheres) is shown. These residues may be part of a common pathway for propagating conformational changes from the ligand-binding pocket to the G-protein coupling domains. Amino acids are numbered using the Ballesteros/Weinstein numbering system, in which the number preceding the dot refers to the transmembrane helix on which an amino acid resides. The second number designates the position relative to the most highly conserved residue among family A GPCRs, numbered 50.
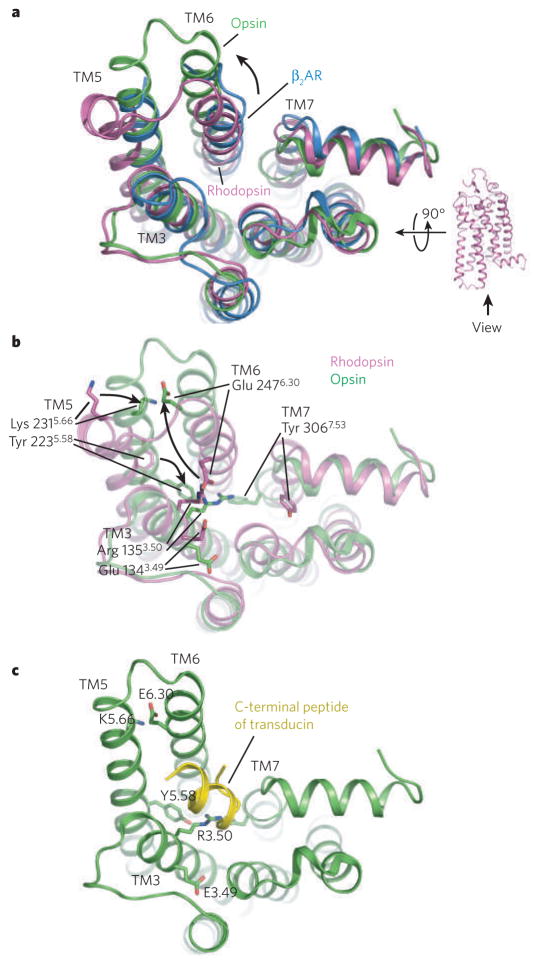
a, A comparison of the cytoplasmic surface of β2AR (blue), rhodopsin (purple) and opsin (green). With the exception of transmembrane segment 5 (TM5), β2AR is more similar to rhodopsin than to opsin. b, Differences between rhosopsin and opsin in interactions between conserved amino acids, including those of the ionic lock. c, Complex between opsin and a peptide representing the carboxyl terminus of the G protein transducin.
References
-
- Fredriksson R, Lagerstrom MC, Lundin LG, Schioth HB. The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Mol Pharmacol. 2003;63:1256–1272. This paper provides a comprehensive analysis of sequence relationships between G-protein-coupled receptors in the human genome. - PubMed
-
- Hoffman BB, Lefkowitz RJ. Adrenergic receptors in the heart. Annu Rev Physiol. 1982;44:475–484. - PubMed
-
- Samama P, Pei G, Costa T, Cotecchia S, Lefkowitz RJ. Negative antagonists promote an inactive conformation of the beta 2-adrenergic receptor. Mol Pharmacol. 1994;45:390–394. - PubMed
-
- Chidiac P, Hebert TE, Valiquette M, Dennis M, Bouvier M. Inverse agonist activity of beta-adrenergic antagonists. Mol Pharmacol. 1994;45:490–499. - PubMed
-
- Xiao RP, Cheng H, Zhou YY, Kuschel M, Lakatta EG. Recent advances in cardiac beta(2)-adrenergic signal transduction. Circ Res. 1999;85:1092–1100. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
