

Review
doi: 10.1038/nature08147.
Emerging roles for lipids in shaping membrane-protein function
Affiliations
- PMID: 19458714
- PMCID: PMC3169427
- DOI: 10.1038/nature08147
Item in Clipboard
Review
Emerging roles for lipids in shaping membrane-protein function
Nature.
.
Abstract
Studies of membrane proteins have revealed a direct link between the lipid environment and the structure and function of some of these proteins. Although some of these effects involve specific chemical interactions between lipids and protein residues, many can be understood in terms of protein-induced perturbations to the membrane shape. The free-energy cost of such perturbations can be estimated quantitatively, and measurements of channel gating in model systems of membrane proteins with their lipid partners are now confirming predictions of simple models.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

a, Ion-channel open probability as a function of pipette pressure for mechanosensitive channels in lipids with different tail lengths. The data are adapted from ref. . The curves are an empirical fit to patch-clamp data using the functional form popen = 1/(1 + exp(−α(P−P1/2)), with the parameters α and P1/2 as fitting parameters. The diagrams on the right show how different tail lengths imply a different hydrophobic mismatch as a result of the boundary conditions at the protein–lipid interface. MscL, mechanosensitive channel of large conductance. PC16, PC18 and PC20 are phospholipid bilayers with lipids with acyl chain lengths of 16, 18 and 20 carbons, respectively. b, Membrane doping and membrane protein function. The diagrams show hypothetical mechanisms whereby the insertion of various molecules can alter the protein–membrane interaction. For example, the asymmetrical insertion of lysolipids in the membrane produces a torque on the protein. The introduction of toxins can alter the boundary conditions between the protein and the surrounding lipids. Finally, small molecules can stiffen the membrane. In principle, all these effects could alter the gating characteristics of a channel.

The two columns correspond to the dilute (proteins do not interact) and crowded (proteins interact) membrane limits. Each column shows the class of geometries found, a diagram of the deformation field in the vicinity of the proteins, and a mathematical description of the energies. For the isolated channel in the dilute limit, the deformation height, u, surrounding a given membrane protein has an elastic decay length, λk, that is smaller than the protein size. The deformation energy around a protein depends on a generic ‘spring constant’, k, determined from membrane properties. The deformation energy scales quadratically with hydrophobic mismatch, uo, and scales approximately linearly with protein circumference, C. For crowded membranes, proteins have a sufficiently small separation distance (d≈λk) that the annulus of deformed material around the proteins overlaps, resulting in an interaction energy that depends on the conformational state, si, of the ith protein.
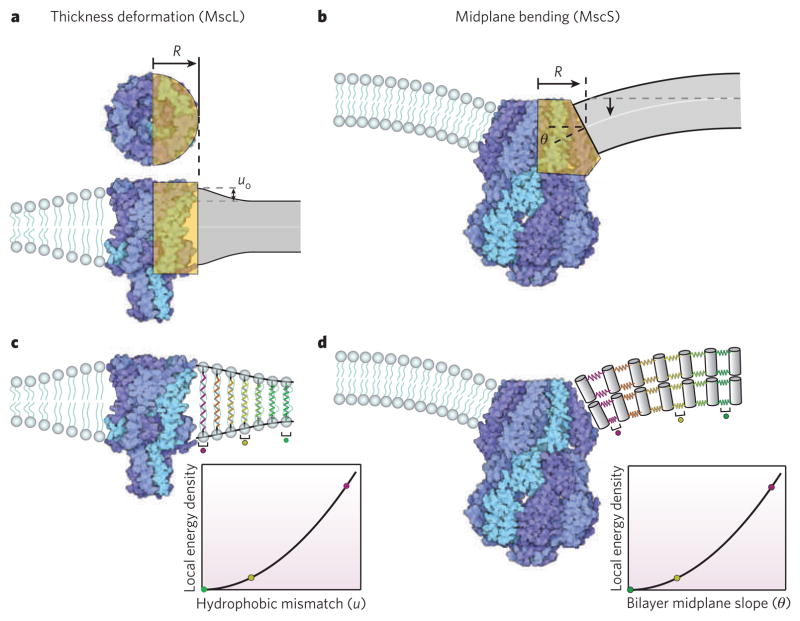
a, Atomic-level structure and an elastic idealization of the mechanosensitive channel of large conductance (MscL) as a rigid cylinder with hydrophobic mismatch at the protein–lipid interface. R, effective radius of channel used in elastic model. uo, hydrophobic mismatch between protein and equilibrium bilayer thickness. b, Atomic-level structure and an elastic idealization of the mechanosensitive channel of small conductance (MscS) as a wedge with a slope that glues continuously onto the surrounding lipids. θ, midplane bending angle at protein–lipid interface. c, Membrane distortion and corresponding free energy of deformation per unit area of membrane surrounding MscL. d, Membrane distortion and corresponding free energy of deformation per unit area of membrane surrounding M33S. In c and d the elastic response of the lipids is captured with springs, and the colour coding indicates the local strain energy density at different distances from the proteins.

Overlap in the deformed membrane between proteins can cause attraction or repulsion over distances comparable to the elastic decay length. The interaction energy between membrane proteins depends on their conformational state and can induce cooperative conformational changes.
References
-
- Engelman DM. Membranes are more mosaic than fluid. Nature. 2005;438:578–580. - PubMed
-
- Perozo E, et al. Physical principles underlying the transduction of bilayer deformation forces during mechanosensitive channel gating. Nature Struct Biol. 2002;9:696–703. This paper describes experiments that show how the gating behaviour of mechanosensitive channels depends on the acyl chain lengths of the lipids around them. - PubMed
-
- Andersen OS, Koeppe RE., II Bilayer thickness and membrane protein function: an energetic perspective. Annu Rev Biophys Biomol Struct. 2007;36:107–130. This useful review provides a variety of examples of the connection between the lipid environment and the function of membrane-bound proteins. - PubMed
-
- Marsh D. Protein modulation of lipids, and vice-versa, in membranes. Biochim Biophys Acta. 2008;1778:1545–1575. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
