

Functional aspects of the solution structure and dynamics of PAF--a highly-stable antifungal protein from Penicillium chrysogenum
- PMID: 19459942
- PMCID: PMC4290664
- DOI: 10.1111/j.1742-4658.2009.07011.x
Functional aspects of the solution structure and dynamics of PAF--a highly-stable antifungal protein from Penicillium chrysogenum
Abstract
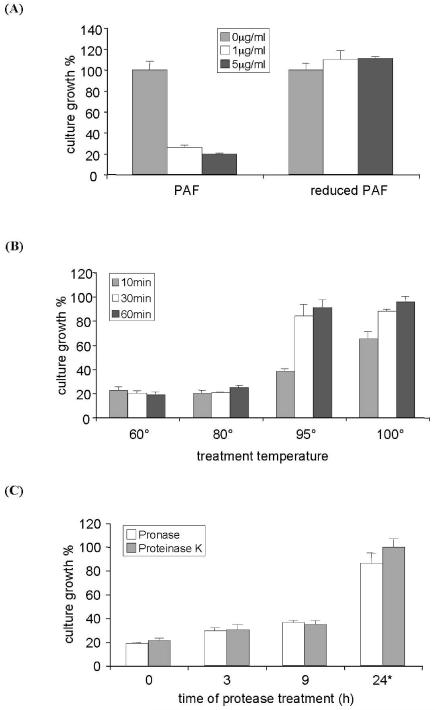
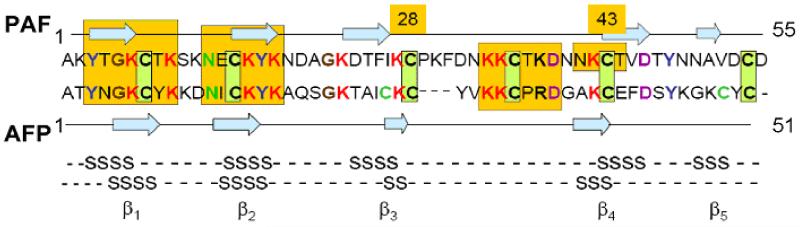
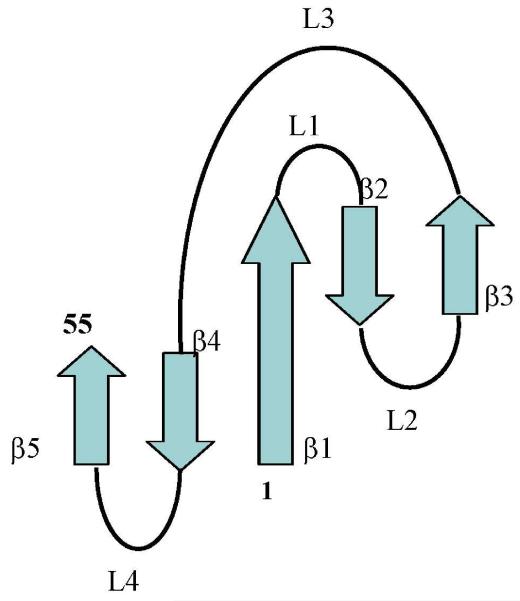
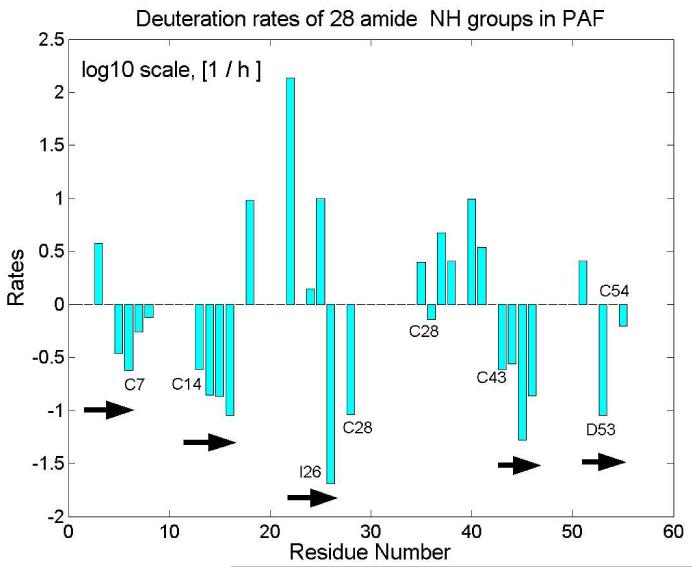
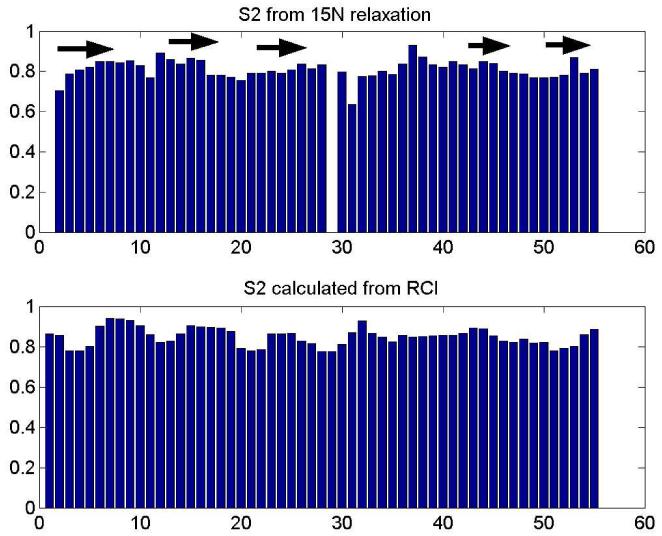
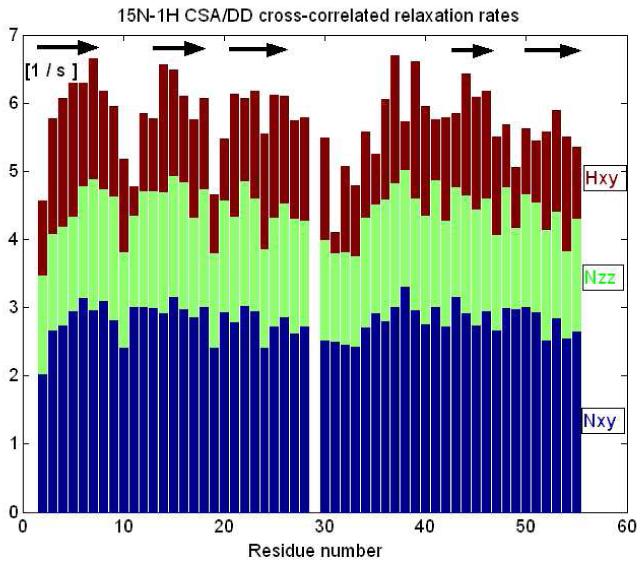
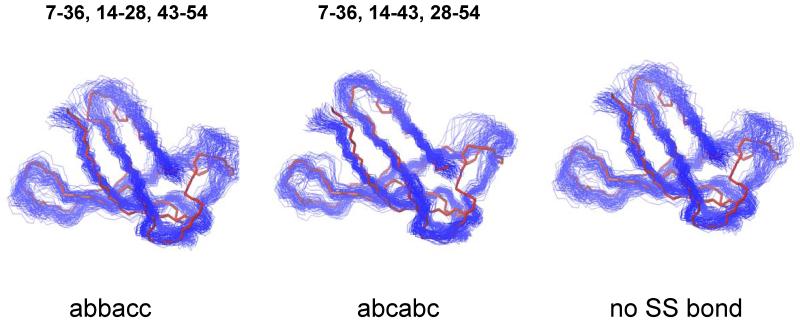
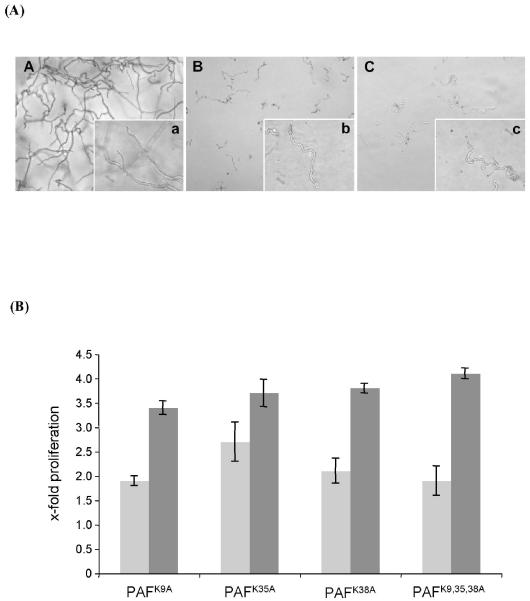
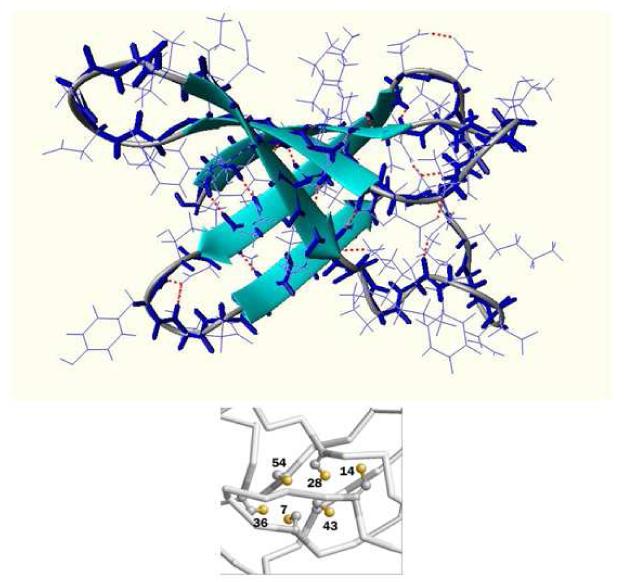
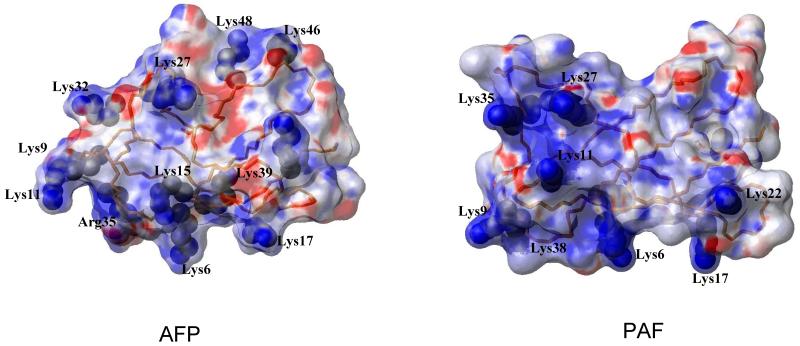
Penicillium antifungal protein (PAF) is a promising antimycotic without toxic effects on mammalian cells and therefore may represent a drug candidate against the often lethal Aspergillus infections that occur in humans. The pathogenesis of PAF on sensitive fungi involves G-protein coupled signalling followed by apoptosis. In the present study, the solution structure of this small, cationic, antifungal protein from Penicillium chrysogenum is determined by NMR. We demonstrate that PAF belongs to the structural classification of proteins fold class of its closest homologue antifungal protein from Aspergillus giganteus. PAF comprises five beta-strands forming two orthogonally packed beta-sheets that share a common interface. The ambiguity in the assignment of two disulfide bonds out of three was investigated by NMR dynamics, together with restrained molecular dynamics calculations. The clue could not be resolved: the two ensembles with different disulfide patterns and the one with no S-S bond exhibit essentially the same fold. (15)N relaxation dispersion and interference experiments did not reveal disulfide bond rearrangements via slow exchange. The measured order parameters and the 3.0 ns correlation time are appropriate for a compact monomeric protein of this size. Using site-directed mutagenesis, we demonstrate that the highly-conserved and positively-charged lysine-rich surface region enhances the toxicity of PAF. However, the binding capability of the oligosaccharide/oligonucleotide binding fold is reduced in PAF compared to antifungal protein as a result of less solvent-exposed aromatic regions, thus explaining the absence of chitobiose binding. The present study lends further support to the understanding of the documented substantial differences between the mode of action of two highly homologous antifungal proteins.
Figures
References
-
- Marx F. Small, basic antifungal proteins secreted from filamentous ascomycetes: a comparative study regarding expression, structure, function and potential application. Applied Microbiology and Biotechnology. 2004;65:133–142. - PubMed
-
- Geisen R. P-nalgiovense carries a gene which is homologous to the paf gene of P-chrysogenum which codes for an antifungal peptide. International Journal of Food Microbiology. 2000;62:95–101. - PubMed
-
- Kaiserer L, Oberparleiter C, Weiler-Gorz R, Burgstaller W, Leiter E, Marx F. Characterization of the Penicillium chrysogenum antifungal protein PAF. Archives of Microbiology. 2003;180:204–210. - PubMed
-
- Lee DG, Shin SY, Maeng CY, Jin ZZ, Kim KL, Hahm KS. Isolation and characterization of a novel antifungal peptide from Aspergillus niger. Biochemical and Biophysical Research Communications. 1999;263:646–651. - PubMed
-
- Marx F, Haas H, Reindl M, Stoffler G, Lottspeich F, Redl B. Cloning, structural organization and regulation of expression of the Penicillium chrysogenum paf gene encoding an abundantly secreted protein with antifungal activity. Gene. 1995;167:167–171. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
