

Genetic architecture of intrinsic antibiotic susceptibility
- PMID: 19462005
- PMCID: PMC2680486
- DOI: 10.1371/journal.pone.0005629
Genetic architecture of intrinsic antibiotic susceptibility
Abstract
Background: Antibiotic exposure rapidly selects for more resistant bacterial strains, and both a drug's chemical structure and a bacterium's cellular network affect the types of mutations acquired.
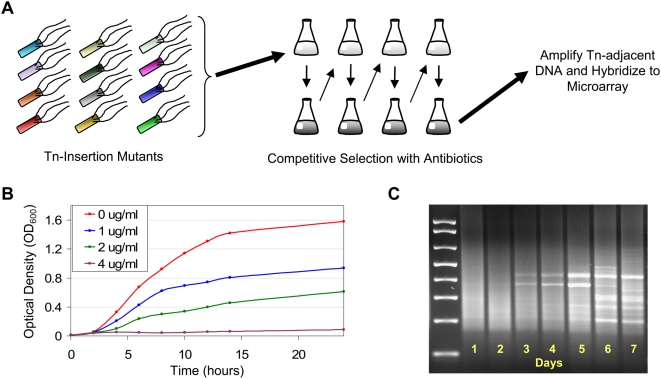
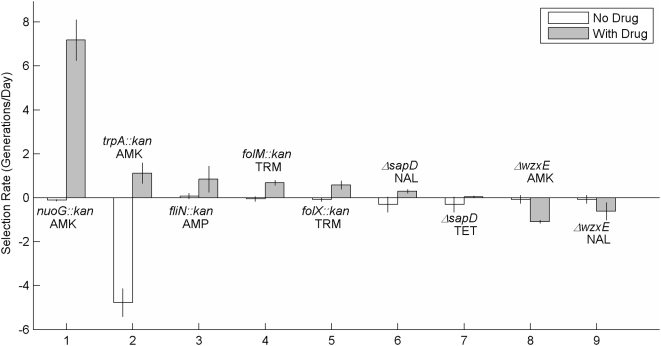
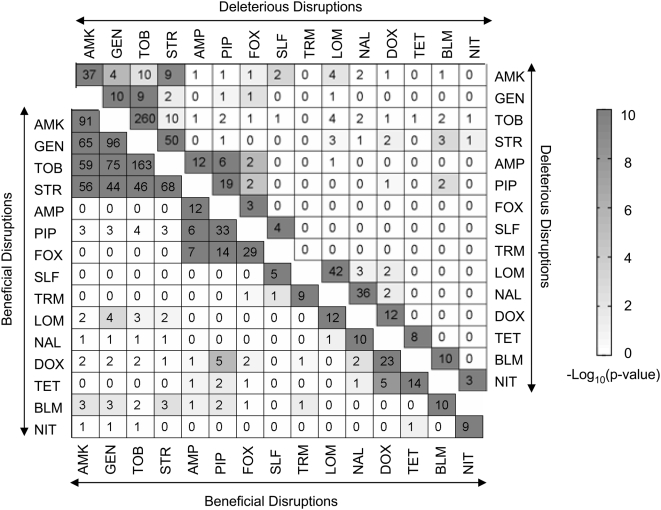
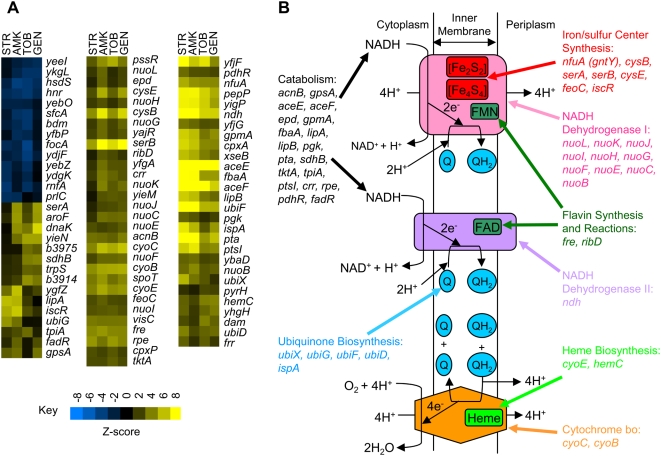
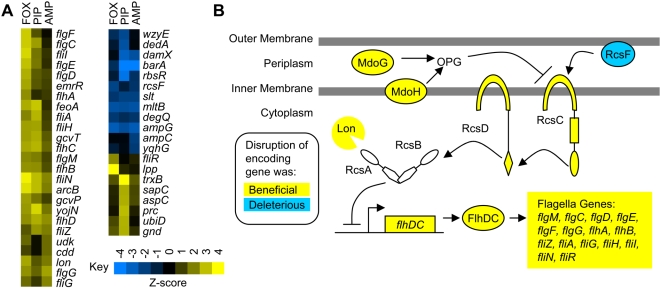
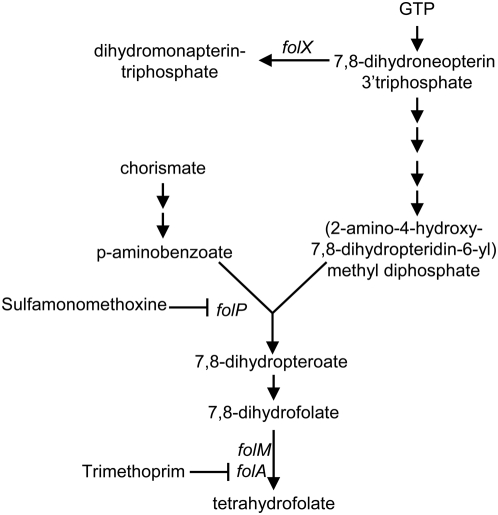
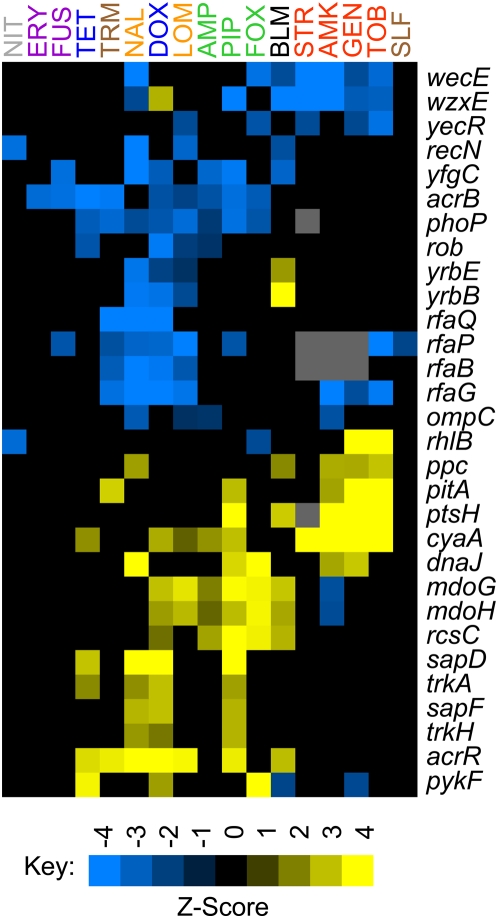
Methodology/principal findings: To better characterize the genetic determinants of antibiotic susceptibility, we exposed a transposon-mutagenized library of Escherichia coli to each of 17 antibiotics that encompass a wide range of drug classes and mechanisms of action. Propagating the library for multiple generations with drug concentrations that moderately inhibited the growth of the isogenic parental strain caused the abundance of strains with even minor fitness advantages or disadvantages to change measurably and reproducibly. Using a microarray-based genetic footprinting strategy, we then determined the quantitative contribution of each gene to E. coli's intrinsic antibiotic susceptibility. We found both loci whose removal increased general antibiotic tolerance as well as pathways whose down-regulation increased tolerance to specific drugs and drug classes. The beneficial mutations identified span multiple pathways, and we identified pairs of mutations that individually provide only minor decreases in antibiotic susceptibility but that combine to provide higher tolerance.
Conclusions/significance: Our results illustrate that a wide-range of mutations can modulate the activity of many cellular resistance processes and demonstrate that E. coli has a large mutational target size for increasing antibiotic tolerance. Furthermore, the work suggests that clinical levels of antibiotic resistance might develop through the sequential accumulation of chromosomal mutations of small individual effect.
Conflict of interest statement
Figures
Similar articles
-
Highly Contingent Phenotypes of Lon Protease Deficiency in Escherichia coli upon Antibiotic Challenge.J Bacteriol. 2020 Jan 15;202(3):e00561-19. doi: 10.1128/JB.00561-19. Print 2020 Jan 15. J Bacteriol. 2020. PMID: 31740490 Free PMC article.
-
Metabolic disruption impairs ribosomal protein levels, resulting in enhanced aminoglycoside tolerance.Elife. 2024 Aug 2;13:RP94903. doi: 10.7554/eLife.94903. Elife. 2024. PMID: 39093940 Free PMC article.
-
Multidrug Intrinsic Resistance Factors in Staphylococcus aureus Identified by Profiling Fitness within High-Diversity Transposon Libraries.mBio. 2016 Aug 16;7(4):e00950-16. doi: 10.1128/mBio.00950-16. mBio. 2016. PMID: 27531908 Free PMC article.
-
The intrinsic resistome of Pseudomonas aeruginosa to β-lactams.Virulence. 2011 Mar-Apr;2(2):144-6. doi: 10.4161/viru.2.2.15014. Epub 2011 Mar 1. Virulence. 2011. PMID: 21304266 Review.
-
Evolutionary Mechanisms Shaping the Maintenance of Antibiotic Resistance.Trends Microbiol. 2018 Aug;26(8):677-691. doi: 10.1016/j.tim.2018.01.005. Epub 2018 Feb 10. Trends Microbiol. 2018. PMID: 29439838 Review.
Cited by
-
Mycoplasmas and Their Antibiotic Resistance: The Problems and Prospects in Controlling Infections.Acta Naturae. 2016 Apr-Jun;8(2):24-34. Acta Naturae. 2016. PMID: 27437137 Free PMC article.
-
The intrinsic resistome of bacterial pathogens.Front Microbiol. 2013 Apr 30;4:103. doi: 10.3389/fmicb.2013.00103. eCollection 2013. Front Microbiol. 2013. PMID: 23641241 Free PMC article.
-
Genotype-phenotype associations in a nonmodel prokaryote.mBio. 2012 Mar 20;3(2):e00001-12. doi: 10.1128/mBio.00001-12. Print 2012. mBio. 2012. PMID: 22434848 Free PMC article.
-
Nonoptimal Gene Expression Creates Latent Potential for Antibiotic Resistance.Mol Biol Evol. 2018 Nov 1;35(11):2669-2684. doi: 10.1093/molbev/msy163. Mol Biol Evol. 2018. PMID: 30169679 Free PMC article.
-
Inactivation of a New Potassium Channel Increases Rifampicin Resistance and Induces Collateral Sensitivity to Hydrophilic Antibiotics in Mycobacterium smegmatis.Antibiotics (Basel). 2022 Apr 12;11(4):509. doi: 10.3390/antibiotics11040509. Antibiotics (Basel). 2022. PMID: 35453260 Free PMC article.
References
-
- Levy SB, Marshall B. Antibacterial resistance worldwide: causes, challenges and responses. Nat Med. 2004;10:S122–129. - PubMed
-
- Walsh C. Where will new antibiotics come from? Nat Rev Microbiol. 2003;1:65–70. - PubMed
-
- Drlica K, Zhao X. Mutant selection window hypothesis updated. Clin Infect Dis. 2007;44:681–688. - PubMed
-
- Davies J, Spiegelman GB, Yim G. The world of subinhibitory antibiotic concentrations. Curr Opin Microbiol. 2006;9:445–453. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
