

Morphine enhances Tat-induced activation in murine microglia
- PMID: 19462331
- PMCID: PMC3121575
- DOI: 10.1080/13550280902913628
Morphine enhances Tat-induced activation in murine microglia
Abstract
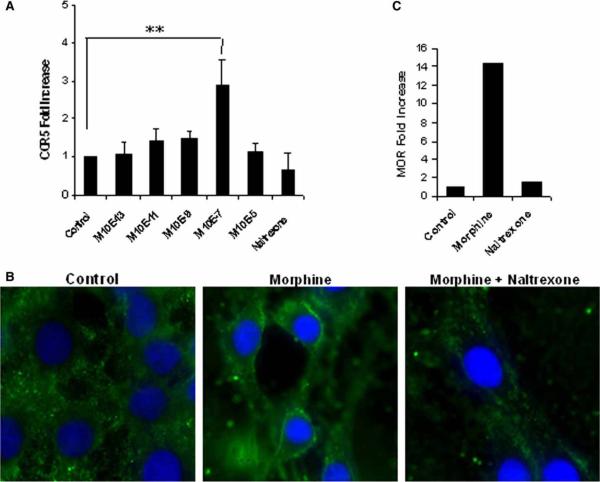
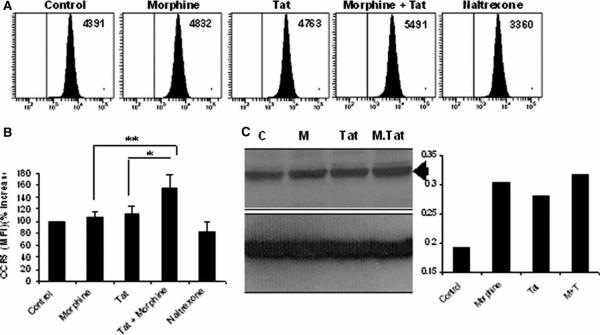
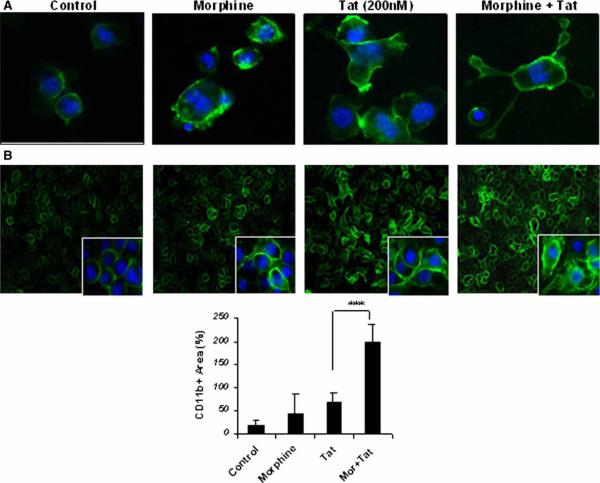
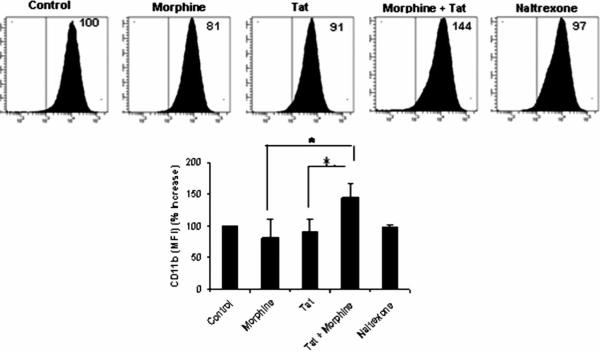
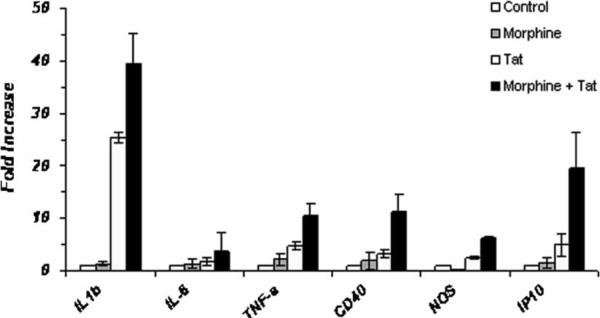
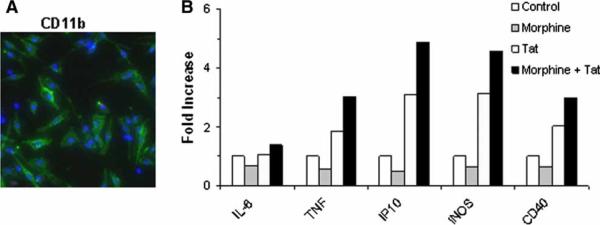
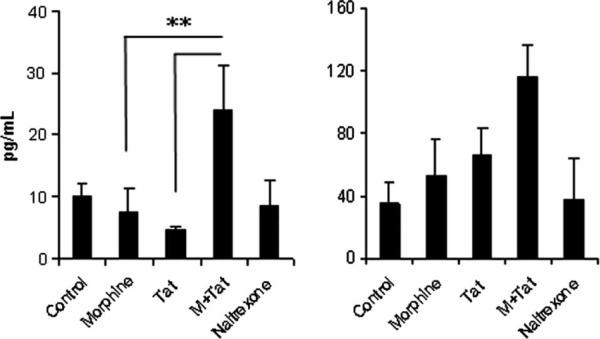
There is increasing evidence that opiates accelerate the pathogenesis and progression of acquired immunodeficiency syndrome (AIDS), as well as the incidence of human immunodeficiency virus (HIV) encephalitis (HIVE), a condition characterized by inflammation, leukocyte infiltration, and microglial activation. The mechanisms, by which the HIV-1 transactivating protein Tat and opioids exacerbate microglial activation, however, are not fully understood. In the current study, we explored the effects of morphine and HIV-1 Tat(1-72) on the activation of mouse BV-2 microglial cells and primary mouse microglia. Both morphine and Tat exposure caused up-regulation of the chemokine receptor CCR5, an effect blocked by the opioid receptor antagonist naltrexone. Morphine in combination with Tat also induced morphological changes in the BV-2 microglia from a quiescent to an activated morphology, with a dramatic increase in the expression of the microglial activation marker CD11b, as compared with cells exposed to either agent alone. In addition, the mRNA expression of inducible nitric oxide synthase (iNOS), CD40 ligand, Interferon-gamma-inducible protein 10 (IP-10), and the proinflammatory cytokines tumor necrosis factor alpha (TNFalpha), interleukin (IL)-1beta, and IL-6, which were elevated with Tat alone, were dramatically enhanced with Tat in the presence of morphine. In summary, these findings shed light on the cooperative effects of morphine and HIV-1 Tat on both microglial activation and HIV coreceptor up-regulation, effects that could result in exacerbated neuropathogenesis.
Figures
Similar articles
-
A central role for glial CCR5 in directing the neuropathological interactions of HIV-1 Tat and opiates.J Neuroinflammation. 2018 Oct 10;15(1):285. doi: 10.1186/s12974-018-1320-4. J Neuroinflammation. 2018. PMID: 30305110 Free PMC article.
-
HIV-1 Tat and opiate-induced changes in astrocytes promote chemotaxis of microglia through the expression of MCP-1 and alternative chemokines.Glia. 2006 Jan 15;53(2):132-46. doi: 10.1002/glia.20262. Glia. 2006. PMID: 16206161 Free PMC article.
-
LRRK2 kinase inhibition prevents pathological microglial phagocytosis in response to HIV-1 Tat protein.J Neuroinflammation. 2012 Nov 29;9:261. doi: 10.1186/1742-2094-9-261. J Neuroinflammation. 2012. PMID: 23190742 Free PMC article.
-
Opiate drug use and the pathophysiology of neuroAIDS.Curr HIV Res. 2012 Jul;10(5):435-52. doi: 10.2174/157016212802138779. Curr HIV Res. 2012. PMID: 22591368 Free PMC article. Review.
-
Crosstalk between components of the blood brain barrier and cells of the CNS in microglial activation in AIDS.Brain Pathol. 2001 Jul;11(3):306-12. doi: 10.1111/j.1750-3639.2001.tb00401.x. Brain Pathol. 2001. PMID: 11414473 Free PMC article. Review.
Cited by
-
Entanglement of CCR5 and Alzheimer's Disease.Front Aging Neurosci. 2019 Aug 7;11:209. doi: 10.3389/fnagi.2019.00209. eCollection 2019. Front Aging Neurosci. 2019. PMID: 31447666 Free PMC article. Review.
-
Increased LPS-induced fever and sickness behavior in adult male and female rats perinatally exposed to morphine.Brain Behav Immun. 2025 May;126:38-52. doi: 10.1016/j.bbi.2025.01.019. Epub 2025 Feb 3. Brain Behav Immun. 2025. PMID: 39909169
-
HIV, opiates, and enteric neuron dysfunction.Neurogastroenterol Motil. 2015 Apr;27(4):449-54. doi: 10.1111/nmo.12539. Neurogastroenterol Motil. 2015. PMID: 25817054 Free PMC article. Review.
-
Modeling the Effects of Morphine on Simian Immunodeficiency Virus Dynamics.PLoS Comput Biol. 2016 Sep 26;12(9):e1005127. doi: 10.1371/journal.pcbi.1005127. eCollection 2016 Sep. PLoS Comput Biol. 2016. PMID: 27668463 Free PMC article.
-
Opioids, microglia, and temporal lobe epilepsy.Front Neurol. 2024 Jan 5;14:1298489. doi: 10.3389/fneur.2023.1298489. eCollection 2023. Front Neurol. 2024. PMID: 38249734 Free PMC article. Review.
References
-
- Adler H, Peterhans E, Jungi TW. Generation and functional characterization of bovine bone marrow-derived macrophages. Vet Immunol Immunopathol. 1994;41:211–227. - PubMed
-
- Alliot F, Godin I, Pessac B. Microglia derive from progenitors, originating from the yolk sac, and which proliferate in the brain. Brain Res Dev Brain Res. 1999;117:145–152. - PubMed
-
- Bell JE, Donaldson YK, Lowrie S, McKenzie CA, Elton RA, Chiswick A, Brettle RP, Ironside JW, Simmonds P. Influence of risk group and zidovudine therapy on the development of HIV encephalitis and cognitive impairment in AIDS patients. AIDS. 1996;10:493–499. - PubMed
-
- Bonwetsch R, Croul S, Richardson MW, Lorenzana C, Del Valle L, Sverstiuk AE, Amini S, Morgello S, Khalili K, Rappaport J. Role of HIV-1 Tat and CC chemokine MIP-1alpha in the pathogenesis of HIV associated central nervous system disorders. J Neuro-Virol. 1999;5:685–694. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
