

Retinoic acid promotes limb induction through effects on body axis extension but is unnecessary for limb patterning
- PMID: 19464179
- PMCID: PMC2701469
- DOI: 10.1016/j.cub.2009.04.059
Retinoic acid promotes limb induction through effects on body axis extension but is unnecessary for limb patterning
Abstract
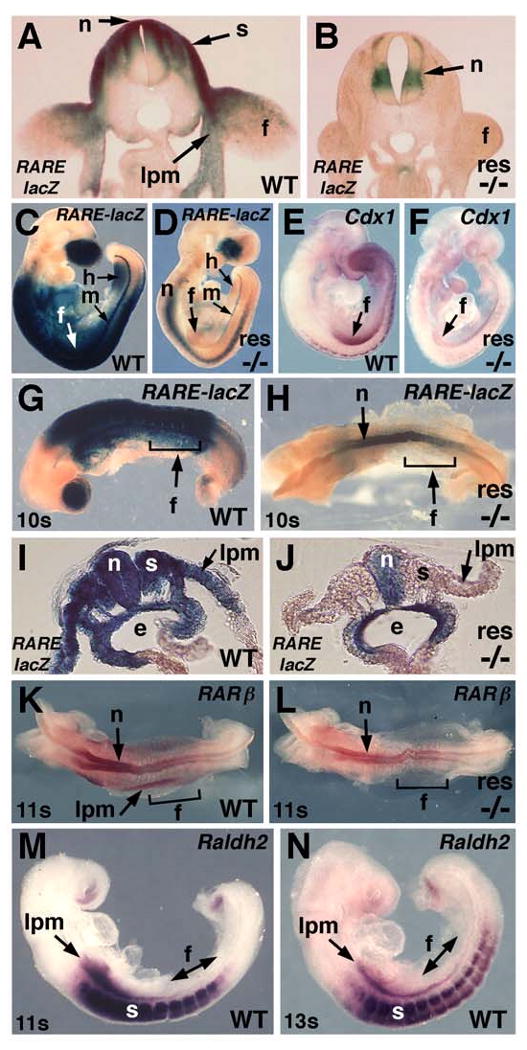
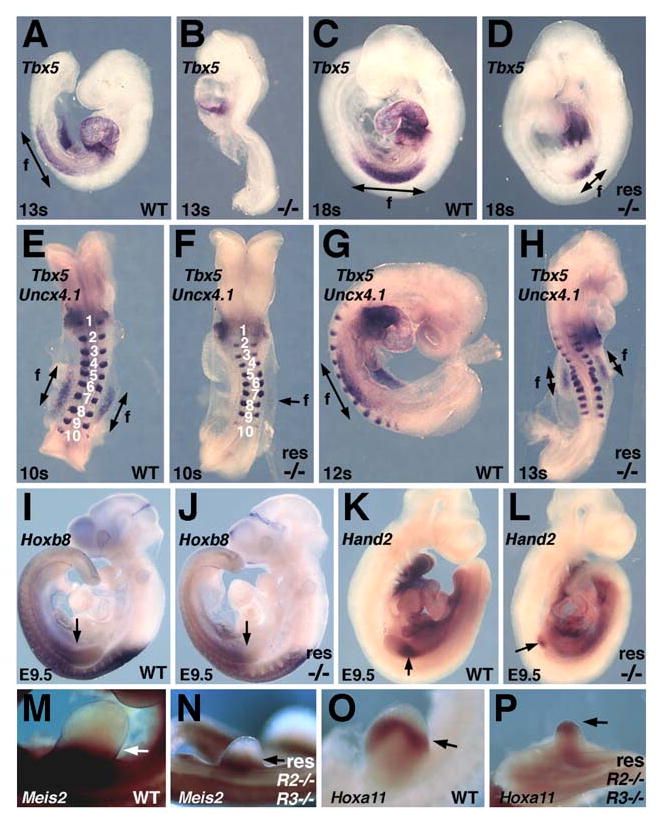
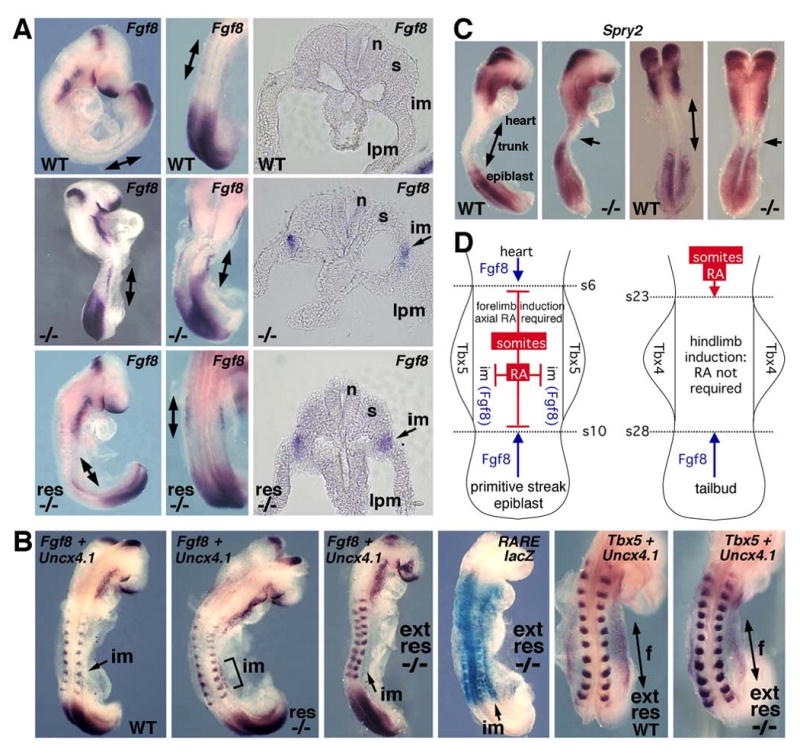
Retinoic acid (RA) is thought to be a key signaling molecule involved in limb bud patterning along the proximodistal or anteroposterior axes functioning through induction of Meis2 and Shh, respectively. Here, we utilize Raldh2-/- and Raldh3-/- mouse embryos lacking RA synthesis to demonstrate that RA signaling is not required for limb expression of Shh and Meis2. We demonstrate that RA action is required outside of the limb field in the body axis during forelimb induction but that RA is unnecessary at later stages when hindlimb budding and patterning occur. We provide evidence for a model of trunk mesodermal RA action in which forelimb induction requires RA repression of Fgf8 in the developing trunk similar to how RA controls somitogenesis and heart development. We demonstrate that pectoral fin development in RA-deficient zebrafish embryos can be rescued by an FGF receptor antagonist SU5402. In addition, embryo ChIP assays demonstrate that RA receptors bind the Fgf8 promoter in vivo. Our findings suggest that RA signaling is not required for limb proximodistal or anteroposterior patterning but that RA inhibition of FGF8 signaling during the early stages of body axis extension provides an environment permissive for induction of forelimb buds.
Figures

Comment in
-
Limb development: the rise and fall of retinoic acid.Curr Biol. 2009 Jul 28;19(14):R558-61. doi: 10.1016/j.cub.2009.06.017. Curr Biol. 2009. PMID: 19640492 Review.
References
-
- Tabin C, Wolpert L. Rethinking the proximodistal axis of the vertebrate limb in the molecular era. Genes & Development. 2007;21:1433–1442. - PubMed
-
- Del Corral RD, Olivera-Martinez I, Goriely A, Gale E, Maden M, Storey K. Opposing FGF and retinoid pathways control ventral neural pattern, neuronal differentiation, and segmentation during body axis extension. Neuron. 2003;40:65–79. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
