

Phylogenetics of Olea (Oleaceae) based on plastid and nuclear ribosomal DNA sequences: tertiary climatic shifts and lineage differentiation times
- PMID: 19465750
- PMCID: PMC2706730
- DOI: 10.1093/aob/mcp105
Phylogenetics of Olea (Oleaceae) based on plastid and nuclear ribosomal DNA sequences: tertiary climatic shifts and lineage differentiation times
Abstract
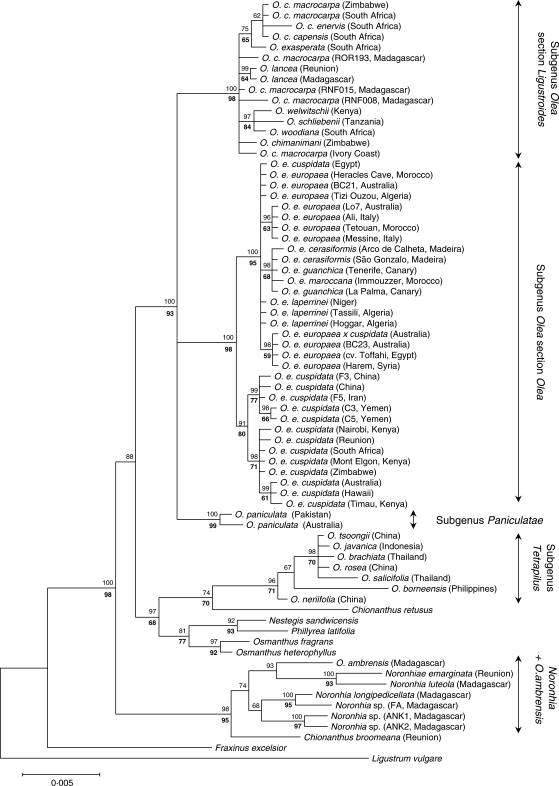
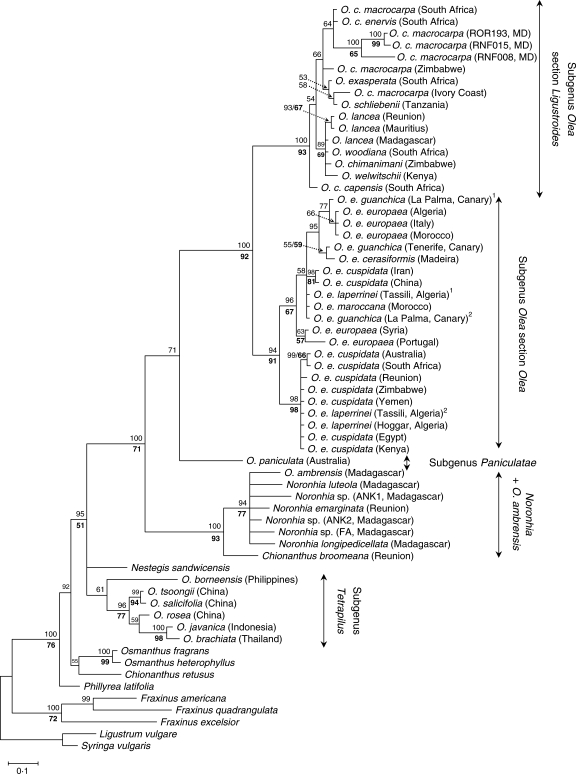
Background and aims: The genus Olea (Oleaceae) includes approx. 40 taxa of evergreen shrubs and trees classified in three subgenera, Olea, Paniculatae and Tetrapilus, the first of which has two sections (Olea and Ligustroides). Olive trees (the O. europaea complex) have been the subject of intensive research, whereas little is known about the phylogenetic relationships among the other species. To clarify the biogeographical history of this group, a molecular analysis of Olea and related genera of Oleaceae is thus necessary.
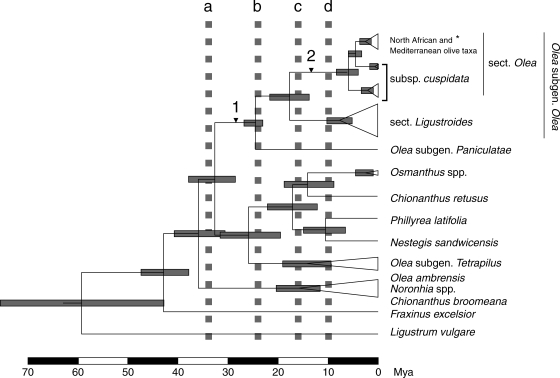
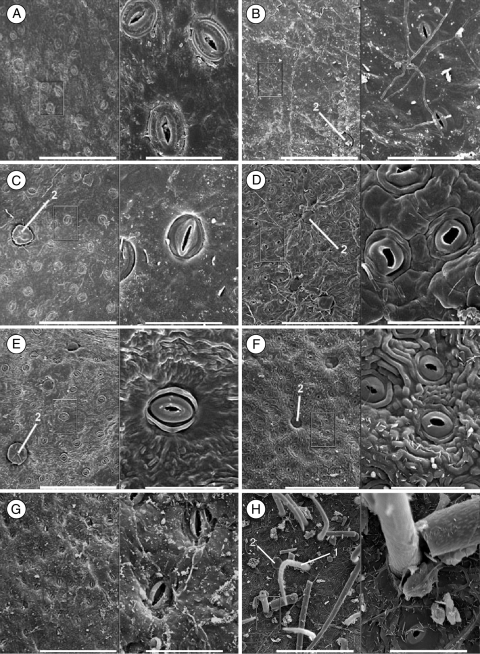
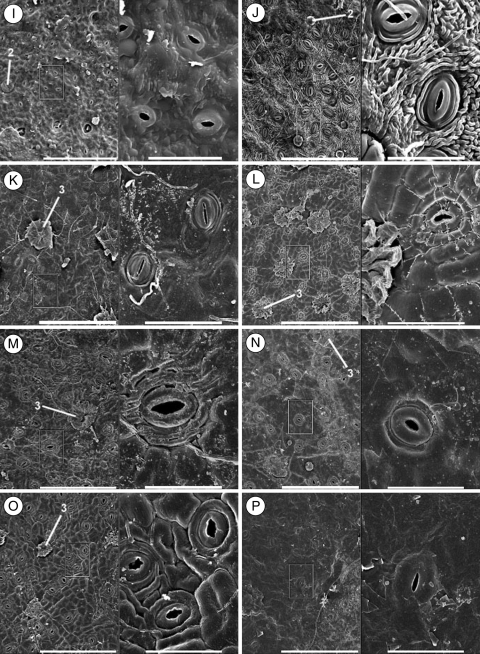
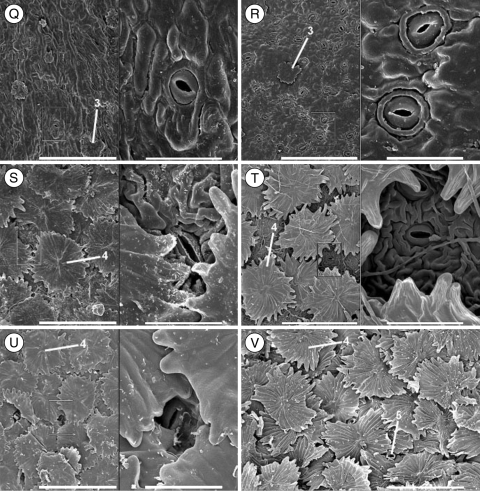
Methods: A phylogeny was built of Olea and related genera based on sequences of the nuclear ribosomal internal transcribed spacer-1 and four plastid regions. Lineage divergence and the evolution of abaxial peltate scales, the latter character linked to drought adaptation, were dated using a Bayesian method.
Key results: Olea is polyphyletic, with O. ambrensis and subgenus Tetrapilus not sharing a most recent common ancestor with the main Olea clade. Partial incongruence between nuclear and plastid phylogenetic reconstructions suggests a reticulation process in the evolution of subgenus Olea. Estimates of divergence times for major groups of Olea during the Tertiary were obtained.
Conclusions: This study indicates the necessity of revising current taxonomic boundaries in Olea. The results also suggest that main lines of evolution were promoted by major Tertiary climatic shifts: (1) the split between subgenera Olea and Paniculatae appears to have taken place at the Miocene-Oligocene boundary; (2) the separation of sections Ligustroides and Olea may have occurred during the Early Miocene following the Mi-1 glaciation; and (3) the diversification within these sections (and the origin of dense abaxial indumentum in section Olea) was concomitant with the aridification of Africa in the Late Miocene.
Figures
References
-
- Álvarez I, Wendel JF. Ribosomal ITS sequences and plant phylogenetic inference. Molecular Phylogenetics and Evolution. 2003;29:417–434. - PubMed
-
- Angiolillo A, Mencuccini M, Baldoni L. Olive genetic diversity assessed using amplified polymorphic fragment length polymorphisms. Theoretical and Applied Genetics. 1999;98:411–421.
-
- Baldoni L, Guerrero C, Sossey-Alaoui K, Abbott AG, Angiolillo A, Lumaret R. Phylogenetic relationships among Olea species based on nucleotide variation at a non-coding chloroplast DNA region. Plant Biology. 2002;4:346–351.
-
- Baldwin BG, Sanderson MJ, Porter JM, Wojciechowski MF, Campbell CS, Donoghue MJ. The ITS region of nuclear ribosomal DNA. A valuable source of evidence on angiosperm phylogeny. Annals of the Missouri Botanical Garden. 1995;82:247–277.
-
- Besnard G, Khadari B, Villemur P, Bervillé A. Cytoplasmic male sterility in the olive (Olea europaea L.) Theoretical and Applied Genetics. 2000;100:1018–1024.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
