

Computational models of the Notch network elucidate mechanisms of context-dependent signaling
- PMID: 19468305
- PMCID: PMC2680760
- DOI: 10.1371/journal.pcbi.1000390
Computational models of the Notch network elucidate mechanisms of context-dependent signaling
Abstract
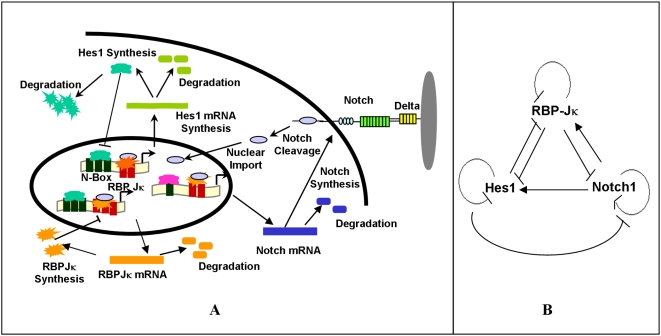
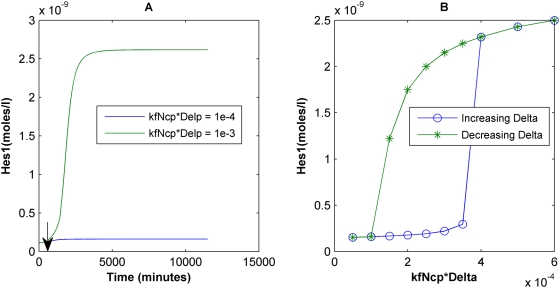
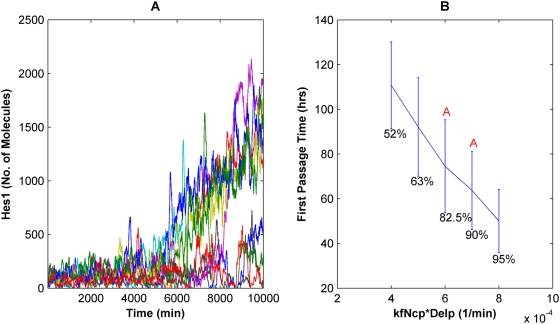
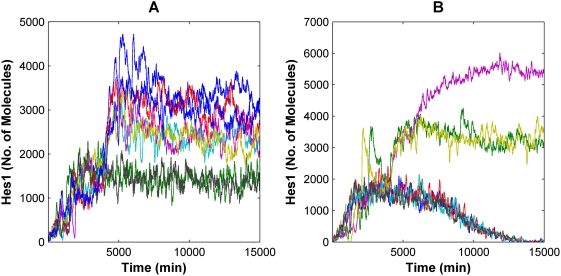
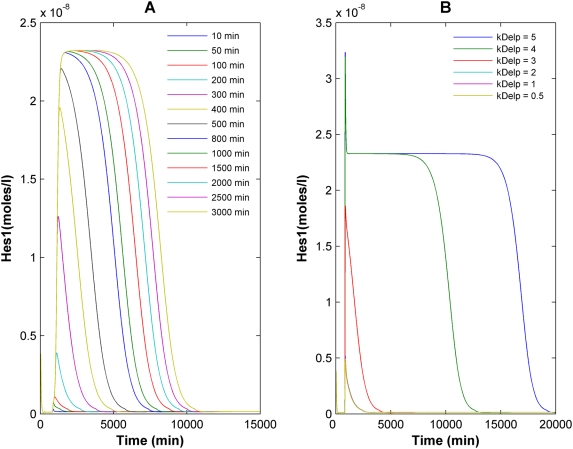
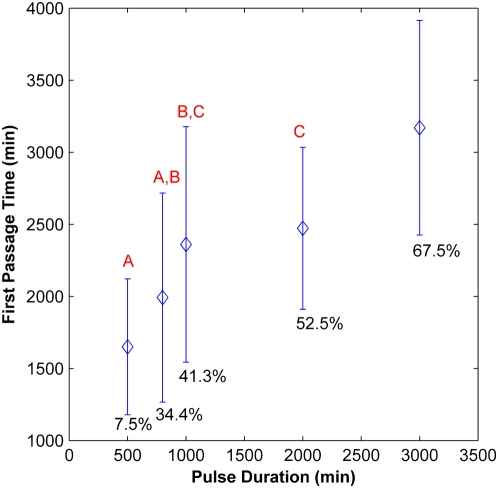
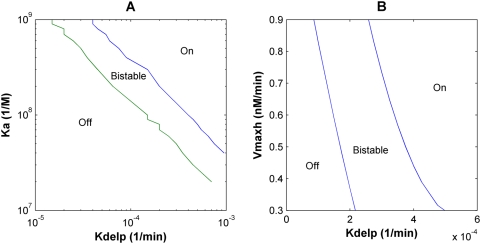
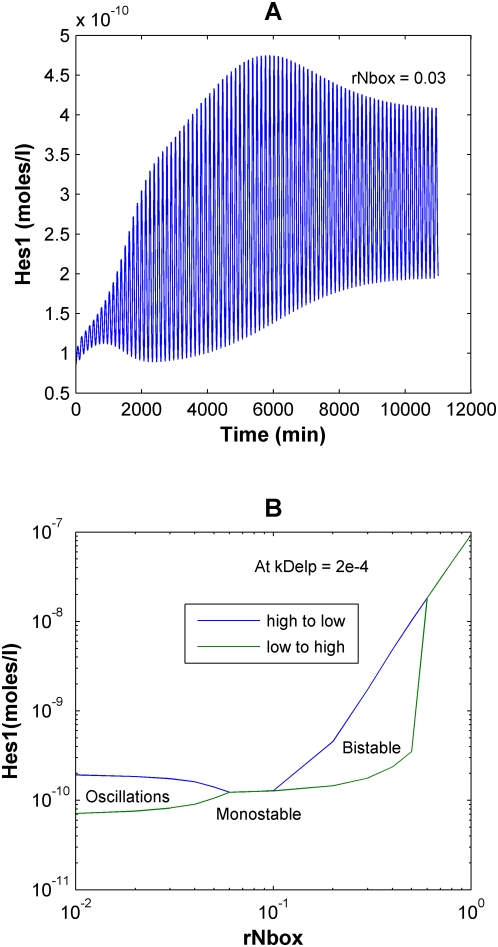
The Notch signaling pathway controls numerous cell fate decisions during development and adulthood through diverse mechanisms. Thus, whereas it functions as an oscillator during somitogenesis, it can mediate an all-or-none cell fate switch to influence pattern formation in various tissues during development. Furthermore, while in some contexts continuous Notch signaling is required, in others a transient Notch signal is sufficient to influence cell fate decisions. However, the signaling mechanisms that underlie these diverse behaviors in different cellular contexts have not been understood. Notch1 along with two downstream transcription factors hes1 and RBP-Jk forms an intricate network of positive and negative feedback loops, and we have implemented a systems biology approach to computationally study this gene regulation network. Our results indicate that the system exhibits bistability and is capable of switching states at a critical level of Notch signaling initiated by its ligand Delta in a particular range of parameter values. In this mode, transient activation of Delta is also capable of inducing prolonged high expression of Hes1, mimicking the "ON" state depending on the intensity and duration of the signal. Furthermore, this system is highly sensitive to certain model parameters and can transition from functioning as a bistable switch to an oscillator by tuning a single parameter value. This parameter, the transcriptional repression constant of hes1, can thus qualitatively govern the behavior of the signaling network. In addition, we find that the system is able to dampen and reduce the effects of biological noise that arise from stochastic effects in gene expression for systems that respond quickly to Notch signaling.This work thus helps our understanding of an important cell fate control system and begins to elucidate how this context dependent signaling system can be modulated in different cellular settings to exhibit entirely different behaviors.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Stochastic simulation of notch signaling reveals novel factors that mediate the differentiation of neural stem cells.J Comput Biol. 2014 Jul;21(7):548-67. doi: 10.1089/cmb.2014.0022. Epub 2014 May 5. J Comput Biol. 2014. PMID: 24798230 Free PMC article.
-
Increased cAMP in monocytes augments Notch signaling mechanisms by elevating RBP-J and transducin-like enhancer of Split (TLE).J Biol Chem. 2013 Jul 26;288(30):21526-36. doi: 10.1074/jbc.M113.465120. Epub 2013 Jun 17. J Biol Chem. 2013. PMID: 23775085 Free PMC article.
-
RUNX3 directly interacts with intracellular domain of Notch1 and suppresses Notch signaling in hepatocellular carcinoma cells.Exp Cell Res. 2010 Jan 15;316(2):149-57. doi: 10.1016/j.yexcr.2009.09.025. Epub 2009 Oct 2. Exp Cell Res. 2010. PMID: 19800882
-
Role of CSL-dependent and independent Notch signaling pathways in cell apoptosis.Apoptosis. 2016 Jan;21(1):1-12. doi: 10.1007/s10495-015-1188-z. Apoptosis. 2016. PMID: 26496776 Review.
-
Bistable Functions and Signaling Motifs in Systems Chemistry: Taking the Next Step Toward Synthetic Cells.Acc Chem Res. 2025 Feb 4;58(3):428-439. doi: 10.1021/acs.accounts.4c00703. Epub 2025 Jan 22. Acc Chem Res. 2025. PMID: 39841921 Free PMC article. Review.
Cited by
-
Delineating cooperative effects of Notch and biomechanical signals on patterned liver differentiation.Commun Biol. 2022 Oct 7;5(1):1073. doi: 10.1038/s42003-022-03840-9. Commun Biol. 2022. PMID: 36207581 Free PMC article.
-
Spatial stochastic modelling of the Hes1 gene regulatory network: intrinsic noise can explain heterogeneity in embryonic stem cell differentiation.J R Soc Interface. 2013 Jan 16;10(80):20120988. doi: 10.1098/rsif.2012.0988. Print 2013 Mar 6. J R Soc Interface. 2013. PMID: 23325756 Free PMC article.
-
Neural stem and progenitor cells in health and disease.Wiley Interdiscip Rev Syst Biol Med. 2013 Nov-Dec;5(6):701-15. doi: 10.1002/wsbm.1239. Epub 2013 Sep 20. Wiley Interdiscip Rev Syst Biol Med. 2013. PMID: 24068527 Free PMC article. Review.
-
Concentration Sensing by the Moving Nucleus in Cell Fate Determination: A Computational Analysis.PLoS One. 2016 Feb 12;11(2):e0149213. doi: 10.1371/journal.pone.0149213. eCollection 2016. PLoS One. 2016. PMID: 26872214 Free PMC article.
-
Stochastic simulation of notch signaling reveals novel factors that mediate the differentiation of neural stem cells.J Comput Biol. 2014 Jul;21(7):548-67. doi: 10.1089/cmb.2014.0022. Epub 2014 May 5. J Comput Biol. 2014. PMID: 24798230 Free PMC article.
References
-
- Keltner NL, Gorman AG. Second messengers. Perspect Psychiatr Care. 2007;43:60–64. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
