

Curcumin modulates efflux mediated by yeast ABC multidrug transporters and is synergistic with antifungals
- PMID: 19470507
- PMCID: PMC2715616
- DOI: 10.1128/AAC.01497-08
Curcumin modulates efflux mediated by yeast ABC multidrug transporters and is synergistic with antifungals
Abstract
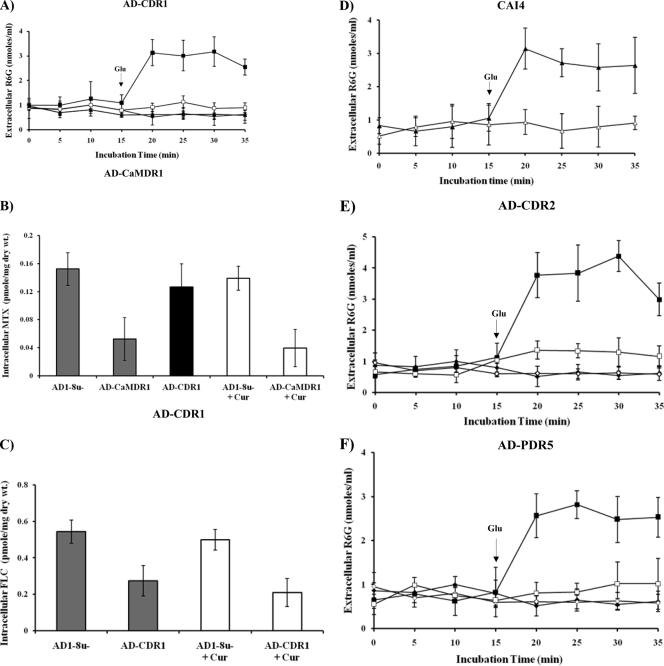
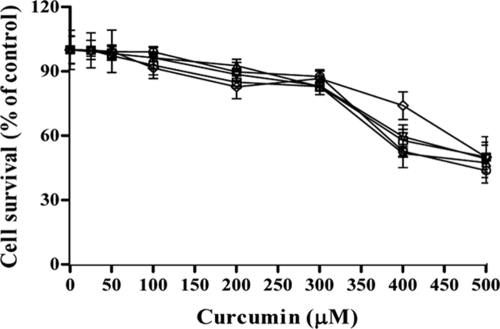
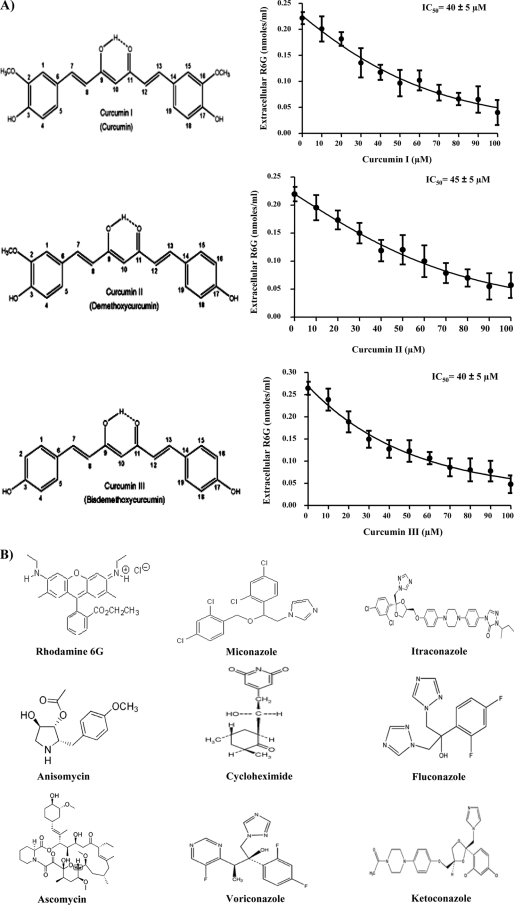
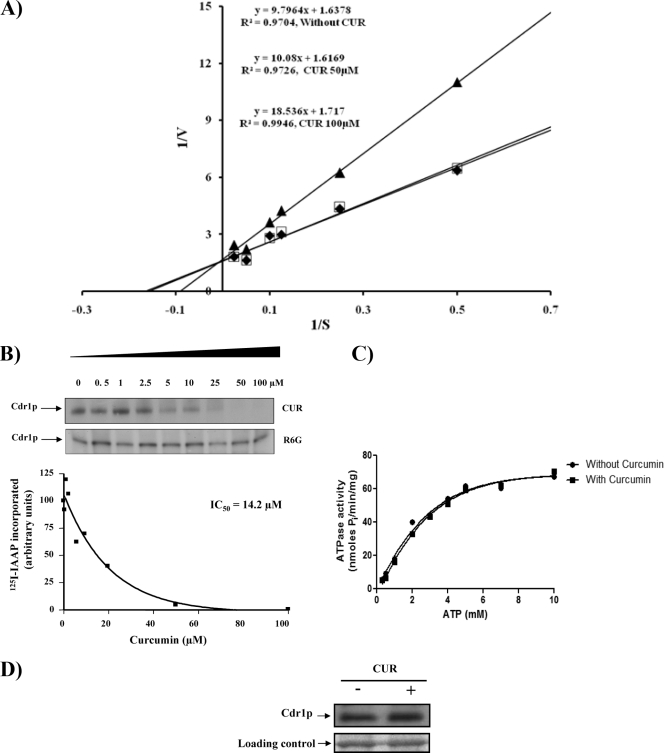
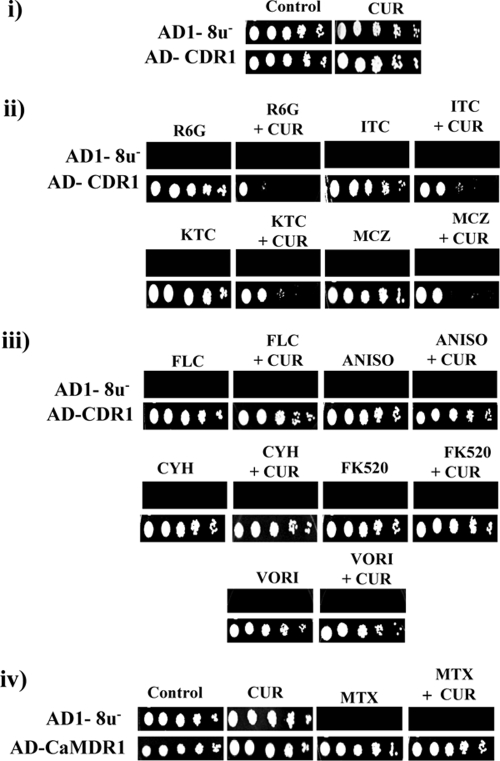
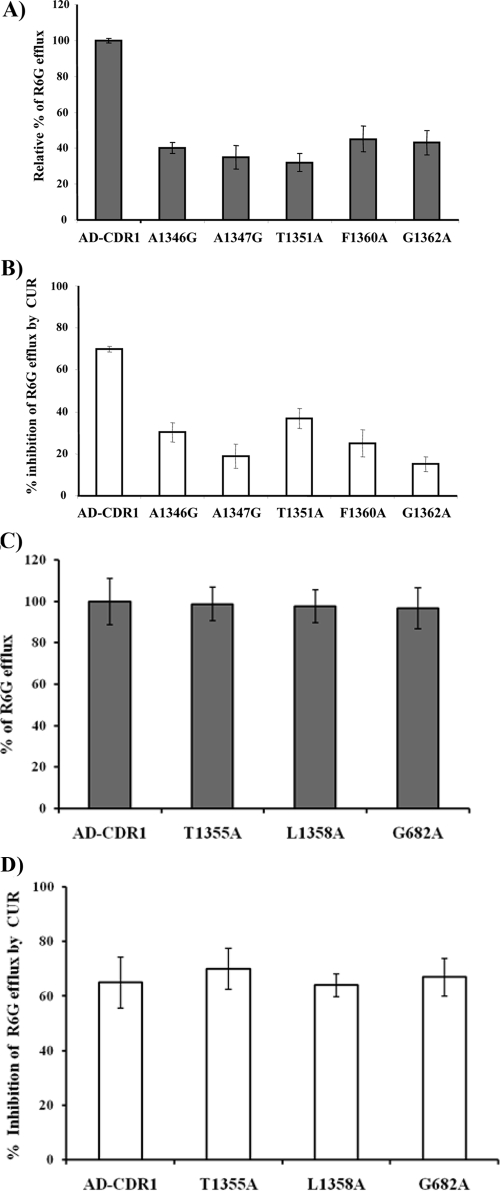
Curcumin (CUR), a natural product of turmeric, from rhizomes of Curcuma longa, is a known agent of reversal of drug resistance phenotypes in cancer cells overexpressing ATP-binding cassette (ABC) transporters, viz., ABCB1, ABCG2, and ABCC1. In the present study, we evaluated whether CUR could also modulate multidrug transporters of yeasts that belong either to the ABC family or to the major facilitator superfamily (MFS). The effect of CUR on multidrug transporter proteins was demonstrated by examining rhodamine 6G (R6G) efflux in Saccharomyces cerevisiae cells overexpressing the Candida albicans ABC transporters Cdr1p and Cdr2p (CaCdr1p and CaCdr2p, respectively) and the MFS transporters CaMdr1p and S. cerevisiae Pdr5p. CUR decreased the extracellular concentration of R6G in ABC transporter-expressing cells but had no effect on methotrexate efflux mediated through the MFS transporter CaMdr1p. CUR competitively inhibited R6G efflux and the photolabeling of CaCdr1p by [(125)I]iodoarylazidoprazosin, a drug analogue of the substrate prazosin (50% inhibitory concentration, 14.2 microM). Notably, the mutant variants of CaCdr1p that displayed abrogated efflux of R6G also showed reduced modulation by CUR. Drug susceptibility testing of ABC protein-expressing cells by spot assays and checkerboard tests revealed that CUR was selectively synergistic with drug substrates such as R6G, ketoconazole, itraconazole, and miconazole but not with fluconazole, voriconazole, anisomycin, cycloheximide, or FK520. Taken together, our results provide the first evidence that CUR modulates only ABC multidrug transporters and could be exploited in combination with certain conventional antifungal drugs to reverse multidrug resistance in Candida cells.
Figures
Similar articles
-
Curcumin potentiates the fungicidal effect of dodecanol by inhibiting drug efflux in wild-type budding yeast.Lett Appl Microbiol. 2019 Jan;68(1):17-23. doi: 10.1111/lam.13083. Epub 2018 Nov 19. Lett Appl Microbiol. 2019. PMID: 30276838
-
The quorum-sensing molecule farnesol is a modulator of drug efflux mediated by ABC multidrug transporters and synergizes with drugs in Candida albicans.Antimicrob Agents Chemother. 2011 Oct;55(10):4834-43. doi: 10.1128/AAC.00344-11. Epub 2011 Jul 18. Antimicrob Agents Chemother. 2011. PMID: 21768514 Free PMC article.
-
The monoamine oxidase A inhibitor clorgyline is a broad-spectrum inhibitor of fungal ABC and MFS transporter efflux pump activities which reverses the azole resistance of Candida albicans and Candida glabrata clinical isolates.Antimicrob Agents Chemother. 2012 Mar;56(3):1508-15. doi: 10.1128/AAC.05706-11. Epub 2011 Dec 27. Antimicrob Agents Chemother. 2012. PMID: 22203607 Free PMC article.
-
Candida Efflux ATPases and Antiporters in Clinical Drug Resistance.Adv Exp Med Biol. 2016;892:351-376. doi: 10.1007/978-3-319-25304-6_15. Adv Exp Med Biol. 2016. PMID: 26721282 Review.
-
Antifungal drug resistance in pathogenic fungi.Med Mycol. 1998;36 Suppl 1:119-28. Med Mycol. 1998. PMID: 9988500 Review.
Cited by
-
Targeting efflux pumps to overcome antifungal drug resistance.Future Med Chem. 2016 Aug;8(12):1485-501. doi: 10.4155/fmc-2016-0050. Epub 2016 Jul 27. Future Med Chem. 2016. PMID: 27463566 Free PMC article. Review.
-
Overcoming Drug Resistance in a Clinical C. albicans Strain Using Photoactivated Curcumin as an Adjuvant.Antibiotics (Basel). 2023 Jul 25;12(8):1230. doi: 10.3390/antibiotics12081230. Antibiotics (Basel). 2023. PMID: 37627652 Free PMC article.
-
Chemosensitization of multidrug resistant Candida albicans by the oxathiolone fused chalcone derivatives.Front Microbiol. 2015 Aug 5;6:783. doi: 10.3389/fmicb.2015.00783. eCollection 2015. Front Microbiol. 2015. PMID: 26300857 Free PMC article.
-
Mechanism of drug resistance in bacteria: efflux pump modulation for designing of new antibiotic enhancers.Folia Microbiol (Praha). 2021 Oct;66(5):727-739. doi: 10.1007/s12223-021-00910-z. Epub 2021 Aug 25. Folia Microbiol (Praha). 2021. PMID: 34431062 Review.
-
Synergistic combinations of antifungals and anti-virulence agents to fight against Candida albicans.Virulence. 2015;6(4):362-71. doi: 10.1080/21505594.2015.1039885. Virulence. 2015. PMID: 26048362 Free PMC article. Review.
References
-
- Ammon, H. P., and M. A. Wahl. 1991. Pharmacology of Curcuma longa. Planta Med. 57:1-7. - PubMed
-
- Anuchapreeda, S., P. Leechanachai, M. M. Smith, S. V. Ambudkar, and P. N. Limtrakul. 2002. Modulation of P-glycoprotein expression and function by curcumin in multidrug-resistant human KB cells. Biochem. Pharmacol. 64:573-582. - PubMed
-
- Chearwae, W., S. Anuchapreeda, K. Nandigama, S. V. Ambudkar, and P. Limtrakul. 2004. Biochemical mechanism of modulation of human P-glycoprotein (ABCB1) by curcumin I, II, and III purified from turmeric powder. Biochem. Pharmacol. 68:2043-2052. - PubMed
-
- Chearwae, W., S. Shukla, P. Limtrakul, and S. V. Ambudkar. 2006. Modulation of the function of the multidrug resistance-linked ATP-binding cassette transporter ABCG2 by the cancer chemopreventive agent curcumin. Mol. Cancer Ther. 5:1995-2006. - PubMed
-
- Chearwae, W., C. P. Wu, H. Y. Chu, T. R. Lee, S. V. Ambudkar, and P. Limtrakul. 2006. Curcuminoids purified from turmeric powder modulate the function of human multidrug resistance protein 1 (ABCC1). Cancer Chemother. Pharmacol. 57:376-388. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Chemical Information
Molecular Biology Databases
