

Maps and streams in the auditory cortex: nonhuman primates illuminate human speech processing
- PMID: 19471271
- PMCID: PMC2846110
- DOI: 10.1038/nn.2331
Maps and streams in the auditory cortex: nonhuman primates illuminate human speech processing
Abstract
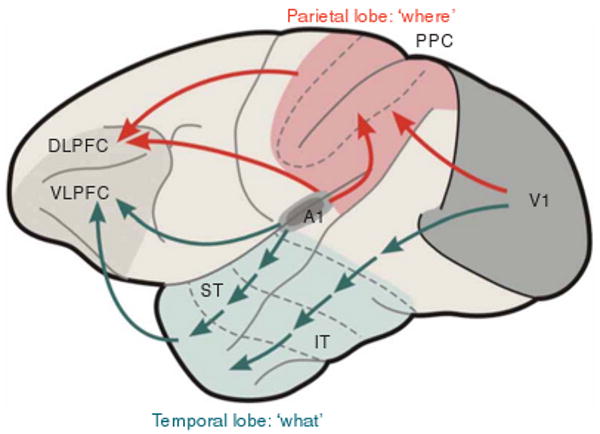
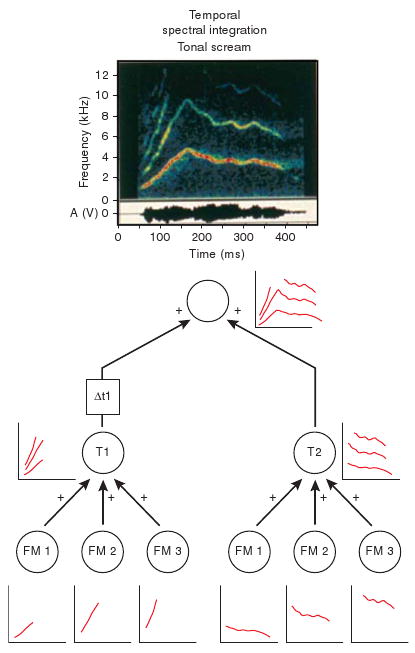
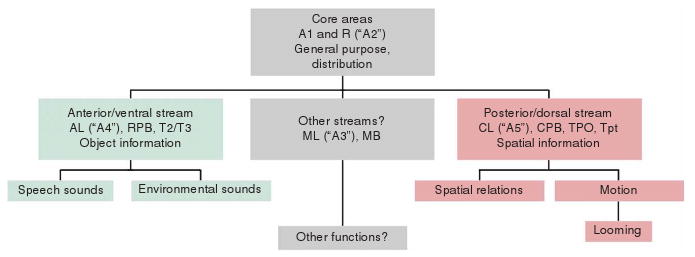
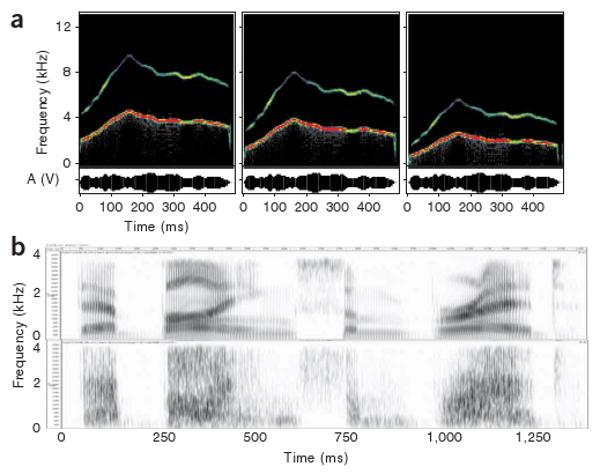
Speech and language are considered uniquely human abilities: animals have communication systems, but they do not match human linguistic skills in terms of recursive structure and combinatorial power. Yet, in evolution, spoken language must have emerged from neural mechanisms at least partially available in animals. In this paper, we will demonstrate how our understanding of speech perception, one important facet of language, has profited from findings and theory in nonhuman primate studies. Chief among these are physiological and anatomical studies showing that primate auditory cortex, across species, shows patterns of hierarchical structure, topographic mapping and streams of functional processing. We will identify roles for different cortical areas in the perceptual processing of speech and review functional imaging work in humans that bears on our understanding of how the brain decodes and monitors speech. A new model connects structures in the temporal, frontal and parietal lobes linking speech perception and production.
Figures
Similar articles
-
The neuroanatomical and functional organization of speech perception.Trends Neurosci. 2003 Feb;26(2):100-7. doi: 10.1016/S0166-2236(02)00037-1. Trends Neurosci. 2003. PMID: 12536133 Review.
-
Neuronal basis of speech comprehension.Hear Res. 2014 Jan;307:121-35. doi: 10.1016/j.heares.2013.09.011. Epub 2013 Oct 7. Hear Res. 2014. PMID: 24113115 Review.
-
Evidence for rapid auditory perception as the foundation of speech processing: a sparse temporal sampling fMRI study.Eur J Neurosci. 2004 Nov;20(9):2447-56. doi: 10.1111/j.1460-9568.2004.03687.x. Eur J Neurosci. 2004. PMID: 15525285
-
Predicting "When" in Discourse Engages the Human Dorsal Auditory Stream: An fMRI Study Using Naturalistic Stories.J Neurosci. 2016 Nov 30;36(48):12180-12191. doi: 10.1523/JNEUROSCI.4100-15.2016. J Neurosci. 2016. PMID: 27903727 Free PMC article.
-
Cortical activity patterns predict speech discrimination ability.Nat Neurosci. 2008 May;11(5):603-8. doi: 10.1038/nn.2109. Epub 2008 Apr 20. Nat Neurosci. 2008. PMID: 18425123 Free PMC article.
Cited by
-
A Bayesian probit model with spatially varying coefficients for brain decoding using fMRI data.Stat Med. 2016 Oct 30;35(24):4380-4397. doi: 10.1002/sim.6999. Epub 2016 May 24. Stat Med. 2016. PMID: 27222305 Free PMC article.
-
Extracting Phonetic Features From Natural Classes: A Mismatch Negativity Study of Mandarin Chinese Retroflex Consonants.Front Hum Neurosci. 2021 Mar 24;15:609898. doi: 10.3389/fnhum.2021.609898. eCollection 2021. Front Hum Neurosci. 2021. PMID: 33841113 Free PMC article.
-
A review and synthesis of the first 20 years of PET and fMRI studies of heard speech, spoken language and reading.Neuroimage. 2012 Aug 15;62(2):816-47. doi: 10.1016/j.neuroimage.2012.04.062. Epub 2012 May 12. Neuroimage. 2012. PMID: 22584224 Free PMC article. Review.
-
The neural control of singing.Front Hum Neurosci. 2013 Jun 3;7:237. doi: 10.3389/fnhum.2013.00237. eCollection 2013. Front Hum Neurosci. 2013. PMID: 23761746 Free PMC article.
-
Auditory properties in the parabelt regions of the superior temporal gyrus in the awake macaque monkey: an initial survey.J Neurosci. 2015 Mar 11;35(10):4140-50. doi: 10.1523/JNEUROSCI.3556-14.2015. J Neurosci. 2015. PMID: 25762661 Free PMC article.
References
-
- Broca P. Remarques sur le siège de la facultè du language articulè: suivies d'une observation d'aphèmie (perte de la parole) Bull Soc Anat Paris. 1861;6:330–357.
-
- Wernicke C. Der aphasische Symptomencomplex: Eine psychologische Studie auf anatomischer Basis. Cohn & Weigert, Breslau; Germany: 1874.
-
- Wise RJ. Language systems in normal and aphasic human subjects: functional imaging studies and inferences from animal studies. Br Med Bull. 2003;65:95–119. - PubMed
-
- Rauschecker JP. Cortical processing of complex sounds. Curr Opin Neurobiol. 1998;8:516–521. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
