

Motif-directed flexible backbone design of functional interactions
- PMID: 19472357
- PMCID: PMC2774439
- DOI: 10.1002/pro.142
Motif-directed flexible backbone design of functional interactions
Abstract
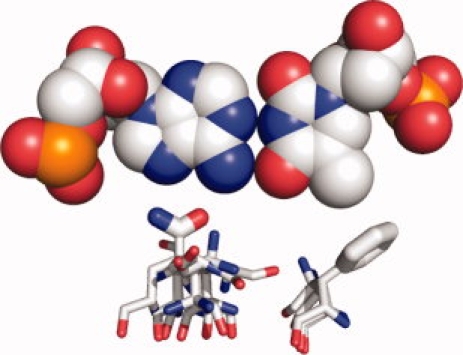
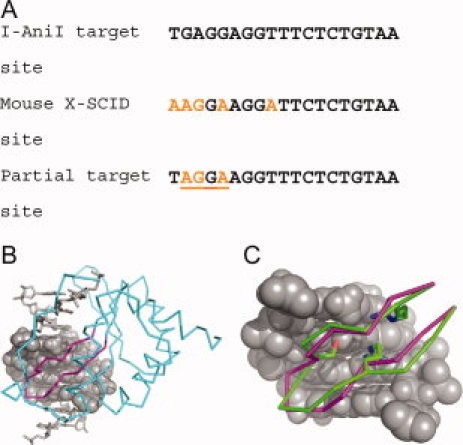
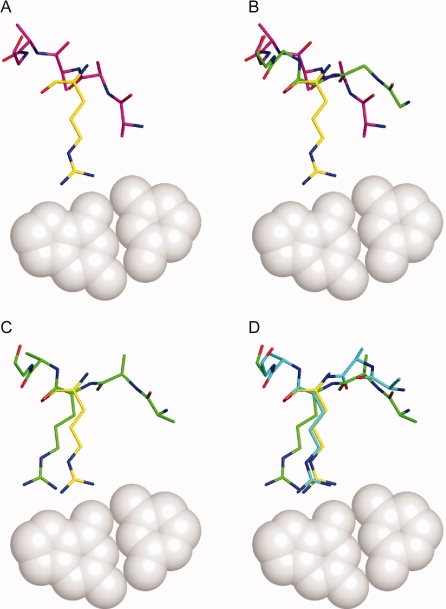
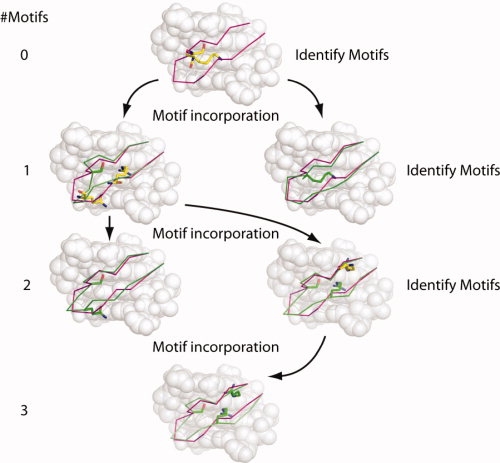
Computational protein design relies on a number of approximations to efficiently search the huge sequence space available to proteins. The fixed backbone and rotamer approximations in particular are important for formulating protein design as a discrete combinatorial optimization problem. However, the resulting coarse-grained sampling of possible side-chain terminal positions is problematic for the design of protein function, which depends on precise positioning of side-chain atoms. Although backbone flexibility can greatly increase the conformation freedom of side-chain functional groups, it is not obvious which backbone movements will generate the critical constellation of atoms responsible for protein function. Here, we report an automated method for identifying protein backbone movements that can give rise to any specified set of desired side-chain atomic placements and interactions, using protein-DNA interfaces as a model system. We use a library of previously observed protein-DNA interactions (motifs) and a rotamer-based description of side-chain conformation freedom to identify placements for the protein backbone that can give rise to a favorable side-chain interaction with DNA. We describe a tree-search algorithm for identifying those combinations of interactions from the library that can be realized with minimal perturbation of the protein backbone. We compare the efficiency of this method with the alternative approach of building and screening alternate backbone conformations.
Figures


Similar articles
-
Improved modeling of side-chain--base interactions and plasticity in protein--DNA interface design.J Mol Biol. 2012 Jun 8;419(3-4):255-74. doi: 10.1016/j.jmb.2012.03.005. Epub 2012 Mar 15. J Mol Biol. 2012. PMID: 22426128 Free PMC article.
-
Motif-directed redesign of enzyme specificity.Protein Sci. 2014 Mar;23(3):312-20. doi: 10.1002/pro.2417. Epub 2014 Feb 4. Protein Sci. 2014. PMID: 24407908 Free PMC article.
-
Dead-end elimination with backbone flexibility.Bioinformatics. 2007 Jul 1;23(13):i185-94. doi: 10.1093/bioinformatics/btm197. Bioinformatics. 2007. PMID: 17646295
-
Principles of flexible protein-protein docking.Proteins. 2008 Nov 1;73(2):271-89. doi: 10.1002/prot.22170. Proteins. 2008. PMID: 18655061 Free PMC article. Review.
-
Macromolecular recognition.Curr Opin Struct Biol. 2005 Apr;15(2):171-5. doi: 10.1016/j.sbi.2005.01.018. Curr Opin Struct Biol. 2005. PMID: 15837175 Review.
Cited by
-
Structure-based modeling of protein: DNA specificity.Brief Funct Genomics. 2015 Jan;14(1):39-49. doi: 10.1093/bfgp/elu044. Epub 2014 Nov 19. Brief Funct Genomics. 2015. PMID: 25414269 Free PMC article. Review.
-
RosettaRemodel: a generalized framework for flexible backbone protein design.PLoS One. 2011;6(8):e24109. doi: 10.1371/journal.pone.0024109. Epub 2011 Aug 31. PLoS One. 2011. PMID: 21909381 Free PMC article.
-
Alteration of enzyme specificity by computational loop remodeling and design.Proc Natl Acad Sci U S A. 2009 Jun 9;106(23):9215-20. doi: 10.1073/pnas.0811070106. Epub 2009 May 21. Proc Natl Acad Sci U S A. 2009. PMID: 19470646 Free PMC article.
-
Proteins comparison through probabilistic optimal structure local alignment.Front Genet. 2014 Sep 2;5:302. doi: 10.3389/fgene.2014.00302. eCollection 2014. Front Genet. 2014. PMID: 25228906 Free PMC article.
-
Reprogramming homing endonuclease specificity through computational design and directed evolution.Nucleic Acids Res. 2014 Feb;42(4):2564-76. doi: 10.1093/nar/gkt1212. Epub 2013 Nov 21. Nucleic Acids Res. 2014. PMID: 24270794 Free PMC article.
References
-
- Pabo C. Molecular technology-designing proteins and peptides. Nature. 1983;301:200–200. - PubMed
-
- Ponder JW, Richards FM. Tertiary templates for proteins. Use of packing criteria in the enumeration of allowed sequences for different structural classes. J Mol Biol. 1987;193:775–791. - PubMed
-
- Harbury PB, Plecs JJ, Tidor B, Alber T, Kim PS. High-resolution protein design with backbone freedom. Science. 1998;282:1462–1467. - PubMed
-
- Kuhlman B, Dantas G, Ireton GC, Varani G, Stoddard BL, Baker D. Design of a novel globular protein fold with atomic-level accuracy. Science. 2003;302:1364–1368. - PubMed
-
- Dahiyat BI, Mayo SL. De novo protein design: fully automated sequence selection. Science. 1997;278:82–87. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
