

Follistatin modulates a BMP autoregulatory loop to control the size and patterning of sensory domains in the developing tongue
- PMID: 19474151
- PMCID: PMC2729339
- DOI: 10.1242/dev.030544
Follistatin modulates a BMP autoregulatory loop to control the size and patterning of sensory domains in the developing tongue
Abstract
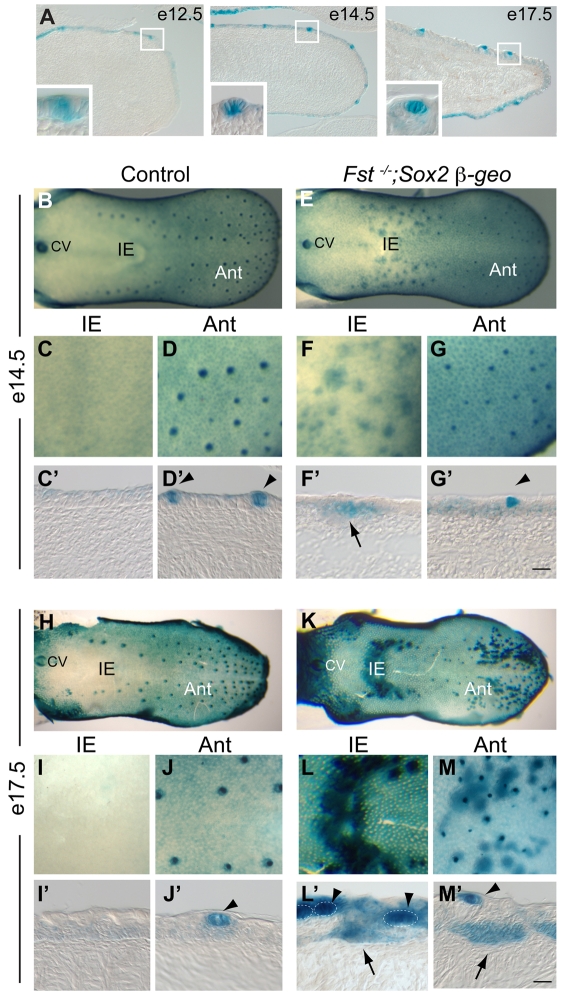
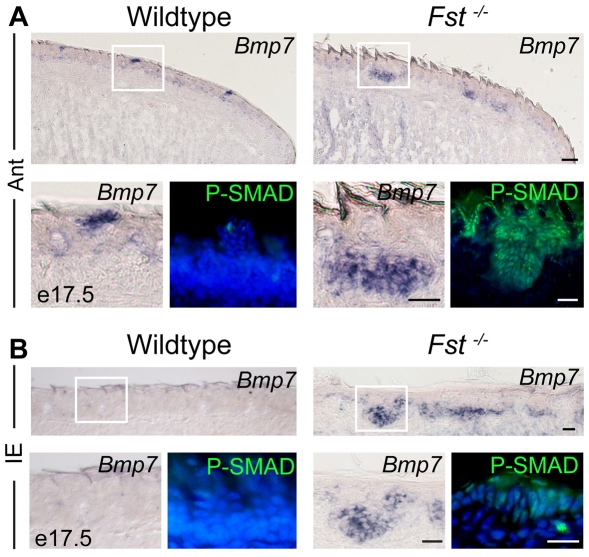
The regenerative capacity of many placode-derived epithelial structures makes them of interest for understanding the molecular control of epithelial stem cells and their niches. Here, we investigate the interaction between the developing epithelium and its surrounding mesenchyme in one such system, the taste papillae and sensory taste buds of the mouse tongue. We identify follistatin (FST) as a mesenchymal factor that controls size, patterning and gustatory cell differentiation in developing taste papillae. FST limits expansion and differentiation of Sox2-expressing taste progenitor cells and negatively regulates the development of taste papillae in the lingual epithelium: in Fst(-/-) tongue, there is both ectopic development of Sox2-expressing taste progenitors and accelerated differentiation of gustatory cells. Loss of Fst leads to elevated activity and increased expression of epithelial Bmp7; the latter effect is consistent with BMP7 positive autoregulation, a phenomenon we demonstrate directly. We show that FST and BMP7 influence the activity and expression of other signaling systems that play important roles in the development of taste papillae and taste buds. In addition, using computational modeling, we show how aberrations in taste papillae patterning in Fst(-/-) mice could result from disruption of an FST-BMP7 regulatory circuit that normally suppresses noise in a process based on diffusion-driven instability. Because inactivation of Bmp7 rescues many of the defects observed in Fst(-/-) tongue, we conclude that interactions between mesenchyme-derived FST and epithelial BMP7 play a central role in the morphogenesis, innervation and maintenance of taste buds and their stem/progenitor cells.
Figures





References
-
- Alappat, S. R., Zhang, Z., Suzuki, K., Zhang, X., Liu, H., Jiang, R., Yamada, G. and Chen, Y. (2005). The cellular and molecular etiology of the cleft secondary palate in Fgf10 mutant mice. Dev. Biol. 277, 102-113. - PubMed
-
- Alonso, L. and Fuchs, E. (2006). The hair cycle. J. Cell Sci. 119, 391-393. - PubMed
-
- Amthor, H., Christ, B., Rashid-Doubell, F., Kemp, C. F., Lang, E. and Patel, K. (2002). Follistatin regulates bone morphogenetic protein-7 (BMP-7) activity to stimulate embryonic muscle growth. Dev. Biol. 243, 115-127. - PubMed
-
- Baker, C. V. and Bronner-Fraser, M. (2001). Vertebrate cranial placodes I. Embryonic induction. Dev. Biol. 232, 1-61. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
