

Excitatory actions of noradrenaline and metabotropic glutamate receptor activation in granule cells of the accessory olfactory bulb
- PMID: 19474170
- PMCID: PMC2724365
- DOI: 10.1152/jn.91093.2008
Excitatory actions of noradrenaline and metabotropic glutamate receptor activation in granule cells of the accessory olfactory bulb
Abstract
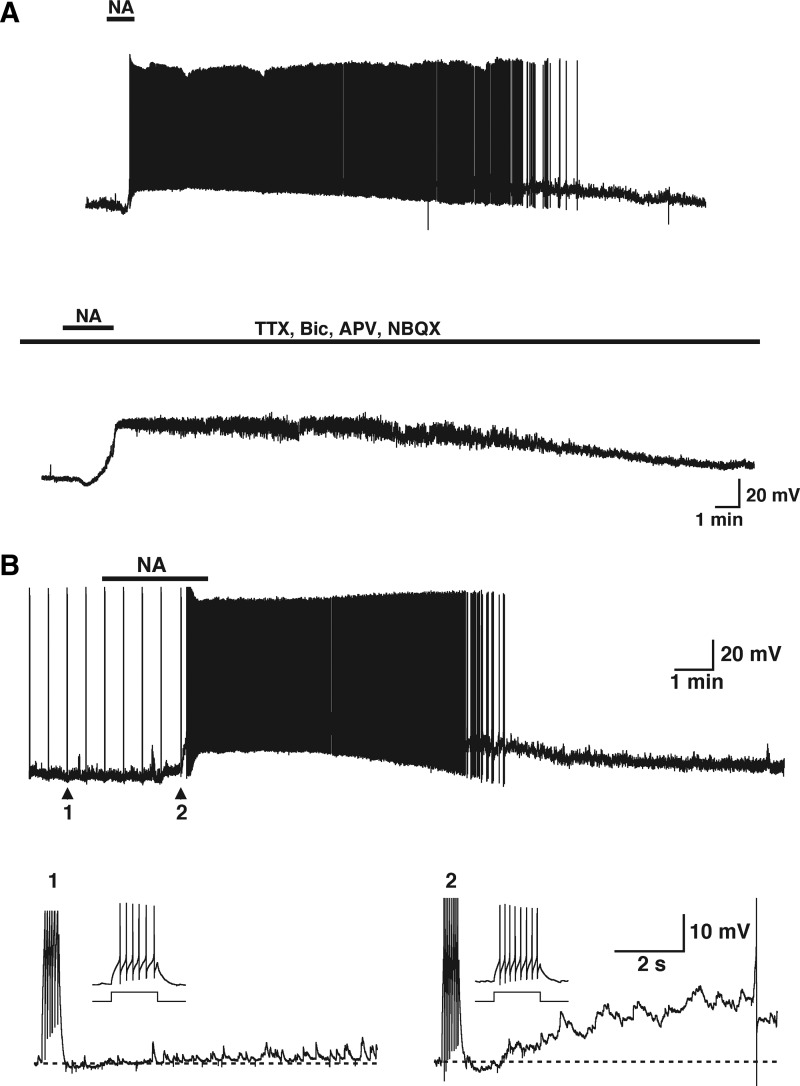
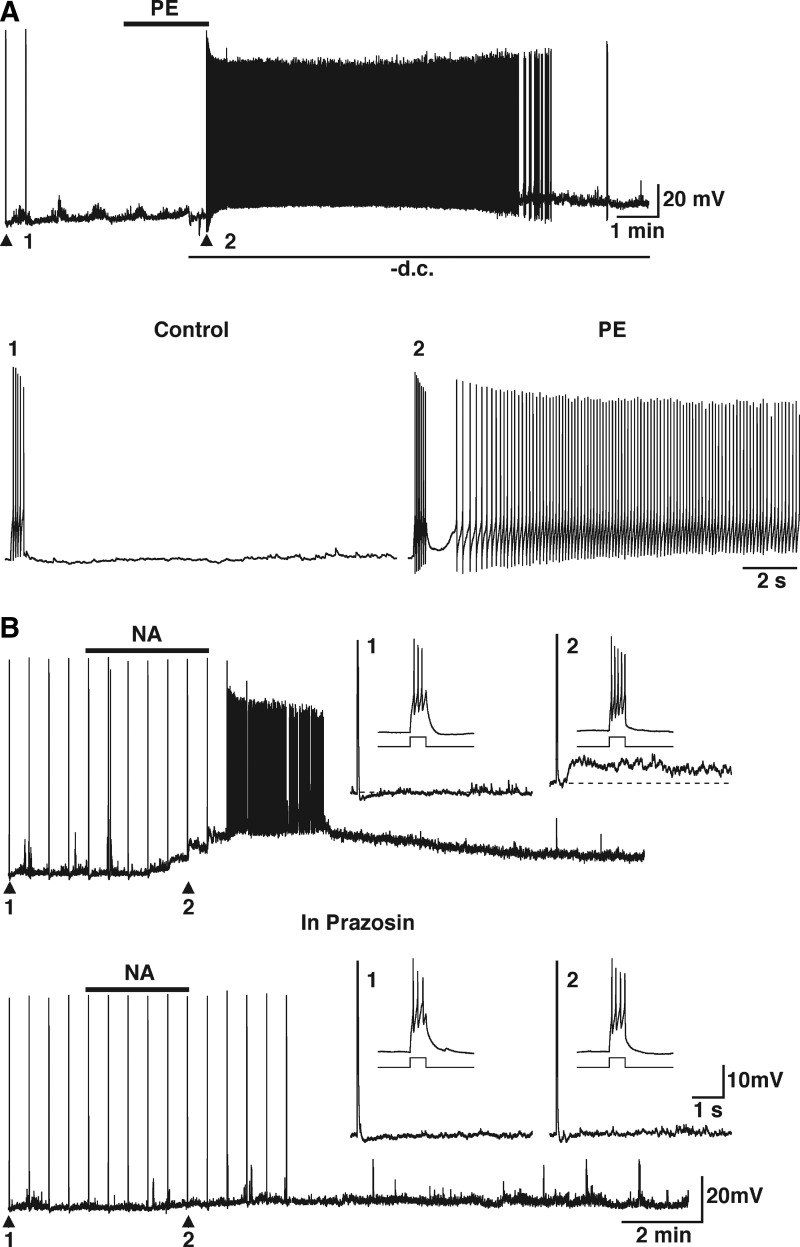
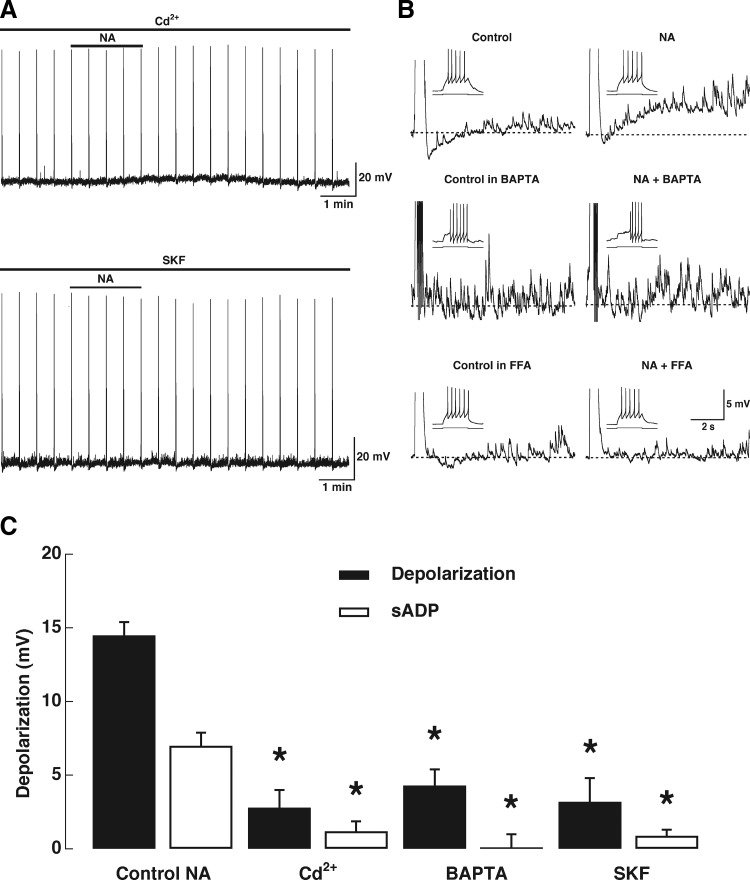
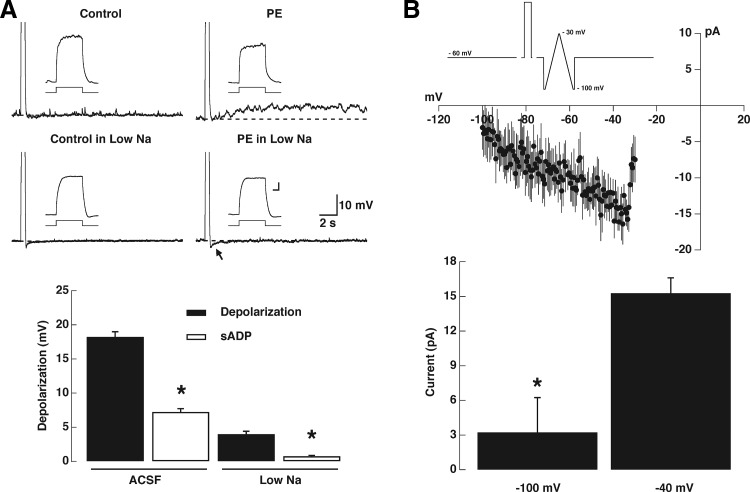
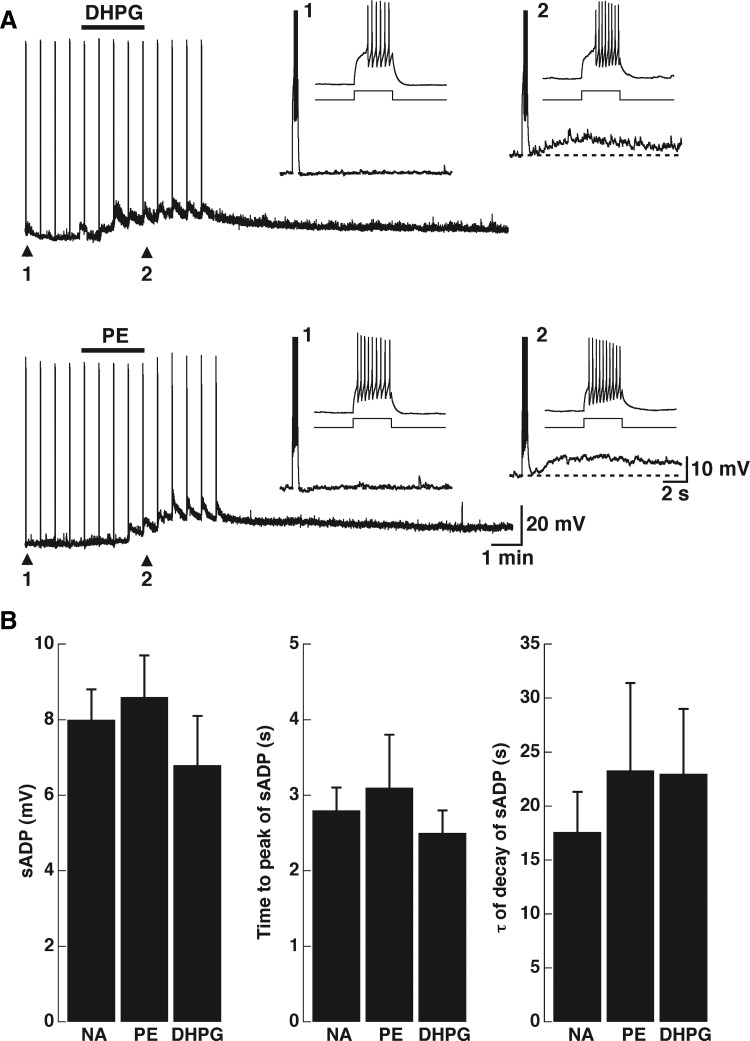
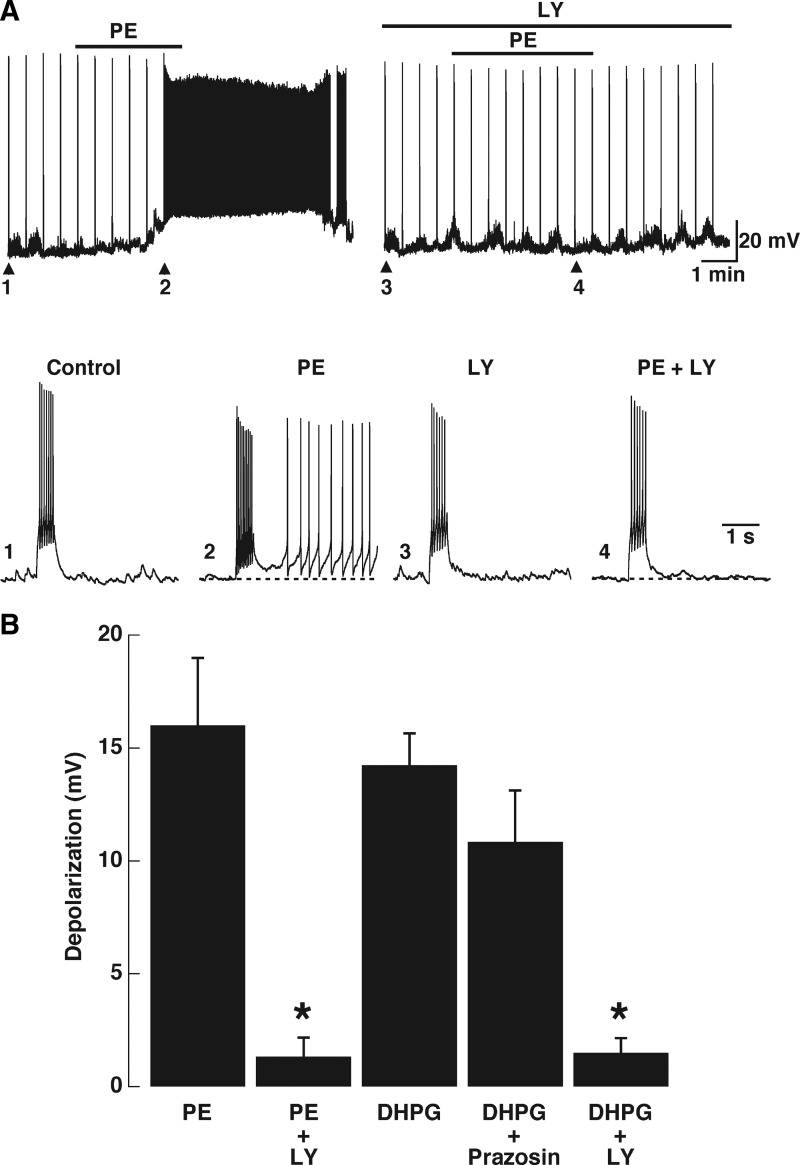
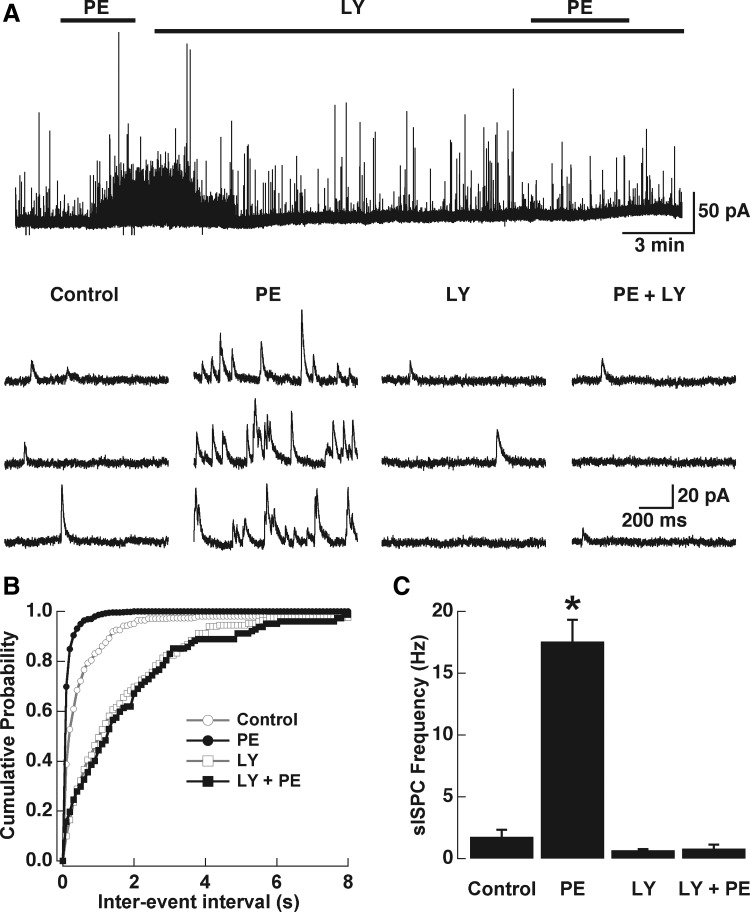
Modulation of dendrodendritic synapses by the noradrenergic system in the accessory olfactory bulb (AOB) plays a key role in the formation of memory in olfactory-mediated behaviors. We have recently shown that noradrenaline (NA) inhibits mitral cells by increasing gamma-aminobutyric acid inhibitory input onto mitral cells in the AOB, suggesting an excitatory action of NA on granule cells (GCs). Here, we show that NA (10 microM) elicits a long-lasting depolarization of GCs. This effect is mediated by activation of alpha(1)-adrenergic receptors as the depolarization is mimicked by phenylephrine (PE, 30 microM) and completely blocked by the alpha(1)-adrenergic receptor antagonist prazosin (300 nM). In addition to this depolarization, application of NA induced the appearance of a slow afterdepolarization (sADP) following a stimulus-elicited train of action potentials. Similarly, the group I metabotropic glutamate receptor (mGluR1) agonist DHPG (10-30 microM) also produced a depolarization of GCs and the appearance of a stimulus-induced sADP. The ionic and voltage dependence and sensitivity to blockers of the sADP suggest that it is mediated by the nonselective cationic conductance I(CAN). Thus the excitatory action resulting from the activation of these receptors could be mediated by a common transduction target. Surprisingly, the excitatory effect of PE on GCs was completely blocked by the mGluR1 antagonist LY367385 (100 microM). Conversely, the effect of DHPG was not antagonized by the alpha(1)-adrenergic receptor antagonist prazosin (300 nM). These results suggest that most of the noradrenergic effect on GCs in the AOB is mediated by potentiation of a basal activity of mGluR1s.
Figures
Similar articles
-
Metabotropic glutamate receptors in the main olfactory bulb drive granule cell-mediated inhibition.J Neurophysiol. 2007 Jan;97(1):858-70. doi: 10.1152/jn.00884.2006. Epub 2006 Nov 8. J Neurophysiol. 2007. PMID: 17093122 Free PMC article.
-
α(1A)-Adrenergic regulation of inhibition in the olfactory bulb.J Physiol. 2013 Apr 1;591(7):1631-43. doi: 10.1113/jphysiol.2012.248591. Epub 2012 Dec 24. J Physiol. 2013. PMID: 23266935 Free PMC article.
-
Olfactory nerve-evoked, metabotropic glutamate receptor-mediated synaptic responses in rat olfactory bulb mitral cells.J Neurophysiol. 2006 Apr;95(4):2233-41. doi: 10.1152/jn.01150.2005. Epub 2006 Jan 4. J Neurophysiol. 2006. PMID: 16394070 Free PMC article.
-
Adrenergic enhancement of inhibitory transmission in the accessory olfactory bulb.J Neurosci. 2006 Mar 22;26(12):3292-8. doi: 10.1523/JNEUROSCI.4768-05.2006. J Neurosci. 2006. PMID: 16554479 Free PMC article.
-
Direct excitation of mitral cells via activation of alpha1-noradrenergic receptors in rat olfactory bulb slices.J Neurophysiol. 2001 Nov;86(5):2173-82. doi: 10.1152/jn.2001.86.5.2173. J Neurophysiol. 2001. PMID: 11698509
Cited by
-
Activation of β-noradrenergic receptors enhances rhythmic bursting in mouse olfactory bulb external tufted cells.J Neurophysiol. 2016 Dec 1;116(6):2604-2614. doi: 10.1152/jn.00034.2016. Epub 2016 Sep 14. J Neurophysiol. 2016. PMID: 27628203 Free PMC article.
-
Differential Muscarinic Modulation in the Olfactory Bulb.J Neurosci. 2015 Jul 29;35(30):10773-85. doi: 10.1523/JNEUROSCI.0099-15.2015. J Neurosci. 2015. PMID: 26224860 Free PMC article.
-
Disruption of centrifugal inhibition to olfactory bulb granule cells impairs olfactory discrimination.Proc Natl Acad Sci U S A. 2013 Sep 3;110(36):14777-82. doi: 10.1073/pnas.1310686110. Epub 2013 Aug 19. Proc Natl Acad Sci U S A. 2013. PMID: 23959889 Free PMC article.
-
In vivo spontaneous activity and coital-evoked inhibition of mouse accessory olfactory bulb output neurons.iScience. 2023 Aug 7;26(9):107545. doi: 10.1016/j.isci.2023.107545. eCollection 2023 Sep 15. iScience. 2023. PMID: 37664596 Free PMC article.
-
Signal Detection and Coding in the Accessory Olfactory System.Chem Senses. 2018 Nov 1;43(9):667-695. doi: 10.1093/chemse/bjy061. Chem Senses. 2018. PMID: 30256909 Free PMC article. Review.
References
-
- Araneda R, Andrade R. 5-Hydroxytryptamine 2 and 5-hydroxytryptamine 1A receptors mediate opposing responses on membrane excitability in rat association cortex. Neuroscience 40: 399–412, 1991. - PubMed
-
- Braga MF, Aroniadou-Anderjaska V, Manion ST, Hough CJ, Li H. Stress impairs alpha(1A) adrenoceptor-mediated noradrenergic facilitation of GABAergic transmission in the basolateral amygdala. Neuropsychopharmacology 29: 45–58, 2004. - PubMed
-
- Brennan PA The nose knows who's who: chemosensory individuality and mate recognition in mice. Horm Behav 46: 231–240, 2004. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources