

Review
doi: 10.1021/cr900097c.
DNA topoisomerase I inhibitors: chemistry, biology, and interfacial inhibition
Affiliations
- PMID: 19476377
- PMCID: PMC2707511
- DOI: 10.1021/cr900097c
Item in Clipboard
Review
DNA topoisomerase I inhibitors: chemistry, biology, and interfacial inhibition
Chem Rev.
2009 Jul.
No abstract available
Figures
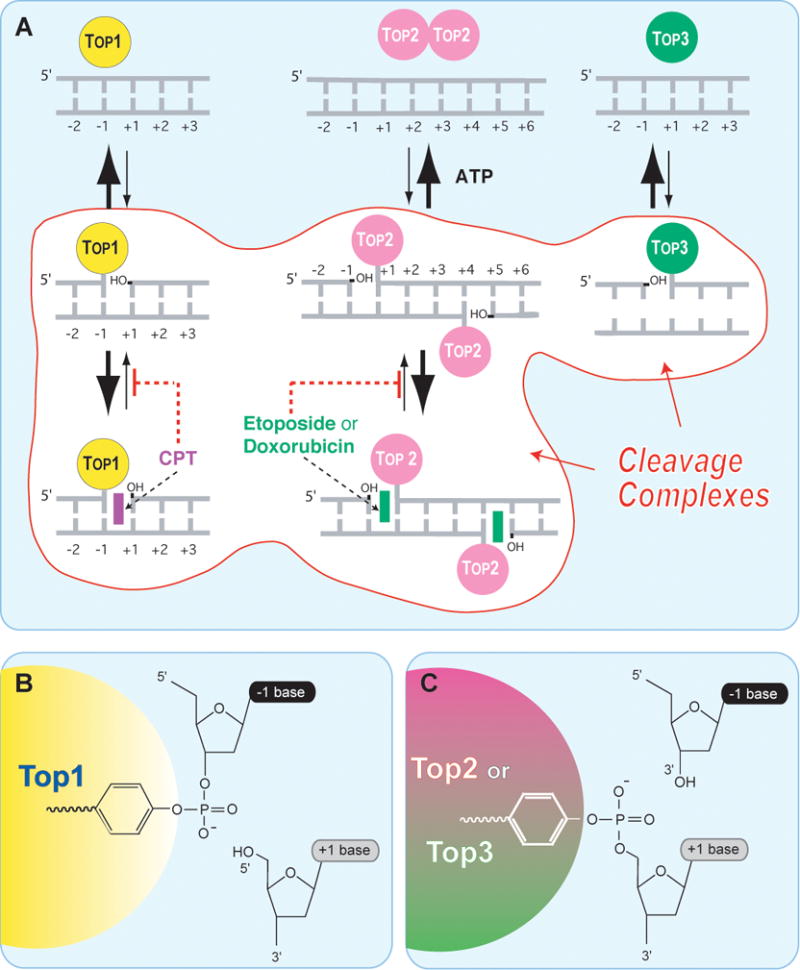
A. Topoisomerases I (Top1 nuclear and Top1mt) bind to double-stranded DNA and form covalent complexes at the 3′-end of the breaks. All other topoisomerases form covalent complexes at the 5′-end of the breaks. Top1 cleavage complexes are selectively stabilized by the natural alkaloid camptothecin (CPT). Topoisomerase II homodimers (Top2α and Top2β) bind to double-stranded DNA and form cleavage complexes with a canonical 4-base pair overhang. Top2 binds and hydrolyze ATP during catalysis. Top2 inhibitors stabilize the Top2 cleavage complexes and are potent anticancer drugs.,,, Topoisomerases III (Top3α and Top3β) bind as monomers to non-canonical DNA structures (single-stranded DNA) in association with a RecQ helicase (BLM in humans, Sgs-1 in budding yeast, Rhq1 in fission yeast). Top3 has been proposed to resolve double-holiday junctions arising from stalled replication forks. Top3 inhibitors have not been reported. B. A topoisomerase catalytic tyrosine residue carries out the nucleophilic attack and breakage of a DNA phosphoester bond. The polarity depends on the topoisomerase type. Topoisomerases I (nuclear and mitochondrial Top1) form a covalent bond with the 3′-DNA end and generate a 5′-hydroxyl-end. This cleavage intermediate (Top1cc) allows controlled rotation of the 5′-end around the intact DNA strand . Under normal conditions, the reaction is reversible. Religation (back arrow from B -> A) is favored over cleavage and requires the alignment of the 5′-hydroxyl-end with the phosphoester tyrosyl-DNA bond for nucleophilic attack. All other human Topo enzymes (Top2 and Top3) have an opposite polarity compared to Top1. They form covalent bonds with the 5′-end of the break and generate 3′-hydroxyl ends.
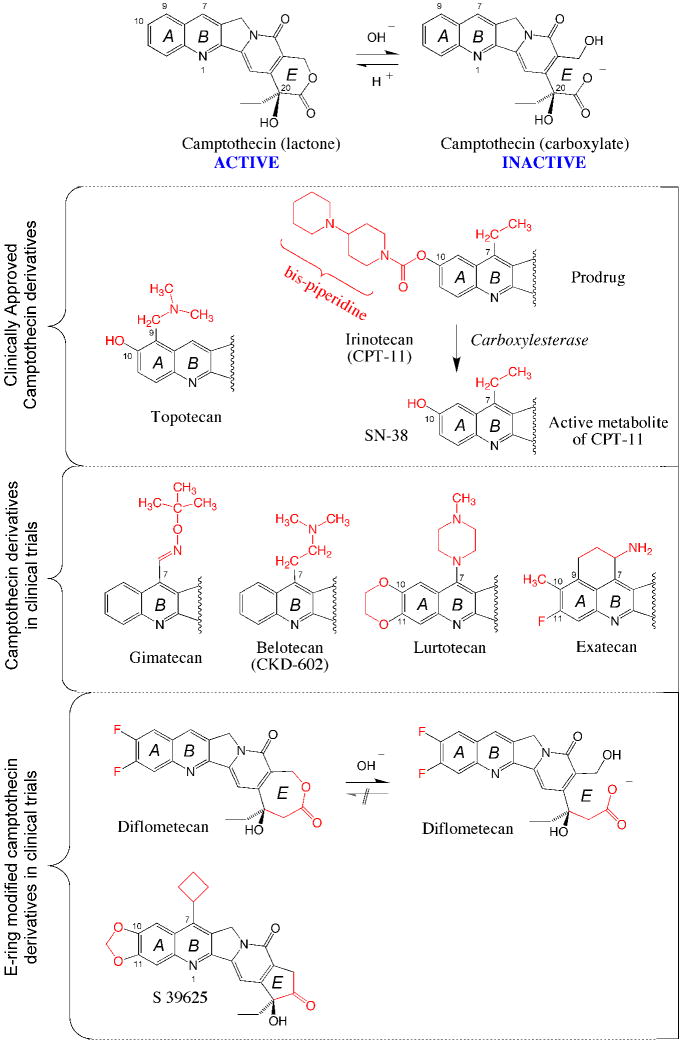
The top scheme shows the equilibrium between the active lactone form and its inactive carboxylate derivative. The lactone is converted into the carboxylate within minutes in human serum at physiological pH . Topotecan (Hycamtin®) and irinotecan are routinely used for IV infusion in cancer treatment. Four CPT derivatives are in clinical trials. Gimatecan (Sigma-Tau®, Novartis®) is an oral derivative developed for the treatment of glioma. Belotecan (CKD602, Camtobell®; Chong Keun Dang Corp.) is a water-soluble derivative given IV. Limited information is available on the ongoing clinical status of lurtotecan and exatecan (Daiichi Pharmaceutical Co Ltd). E-ring modifications have been introduced to generate synthetic analogs with limited (but irreversible) E-ring opening (Diflometecan; Beaufour-Ipsen) ,,, and no ring opening (S 39625; Servier) ,,. Both E-ring-modified derivatives are given IV.
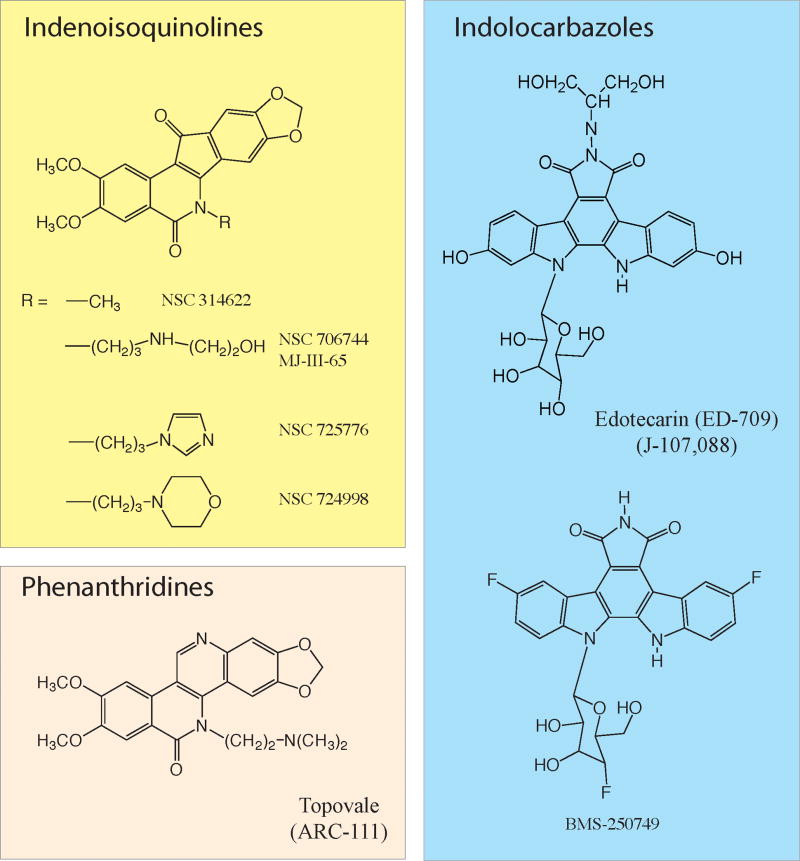
Three chemical families are described (for further information see 52). Two indenoisoquinolines (NSC 725776 and NSC 724998) are in preclinical development at the NCI (Joint patent NCI-Purdue University). ARC-111 is a phenanthridine derivative licensed to Genzyme Co. The indolocarbazoles are further have been tested in clinical trials. Information is limited on their ongoing clinical development.

(A) Top1 is mostly associated non-covalently with chromatin. (B) Top1 relaxes DNA by making single-strand breaks that are generated by the covalent linkage of Top1 to the 3′-end of DNA (Top1cc; see Figure 1). (C) Camptothecins (see Figure 2) or non-camptothecin Top1 inhibitors (see Figure 3) bind reversibly to the Top1cc and slow down DNA religation. (D) Ternary complex including Top1 (yellow), DNA (dark blue ribbons), and an indenoisoquinoline or CPT (green and red in the middle).,,, (E) Same structure except Top1 is in ribbon representation. (F) Hydrogen bond network between camptothecin and Top1 amino acid residues. (G). Hydrogen bond network between the indenoisoquinoline derivative MJ-238 and Top1. Note that mutation of asparagine 722 to serine (N722S), which confers resistance to camptothecin and only partially to indenoisoquinolines, is also present in camptothecin-producing plants.
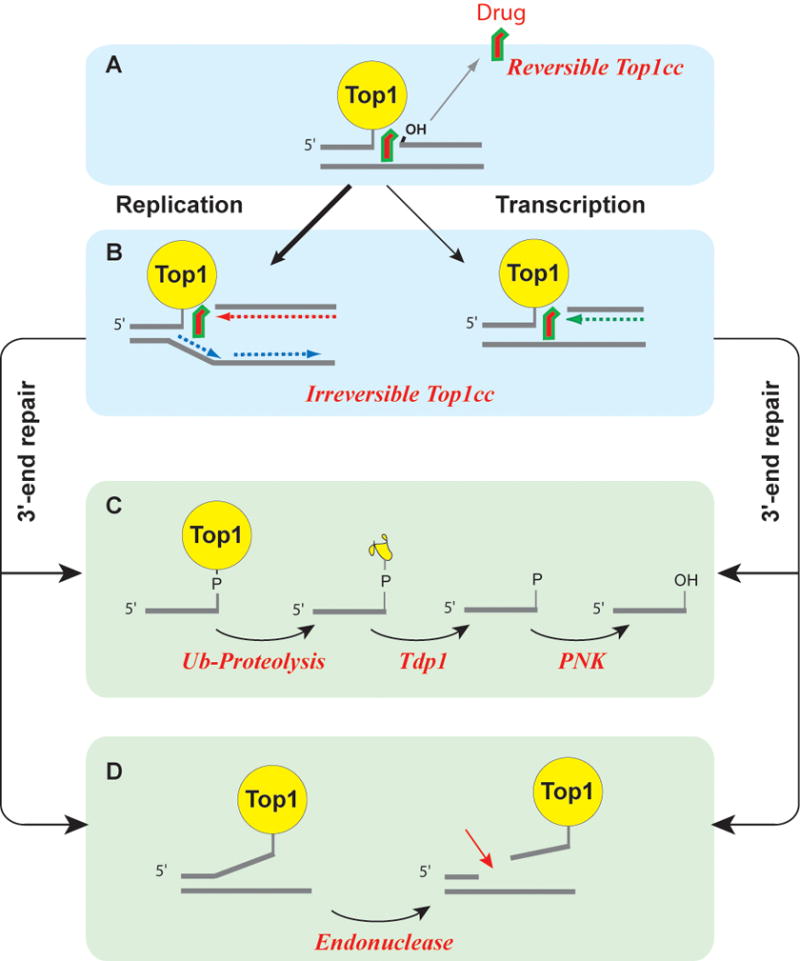
(A) Most Top1cc are reversible as Top1 inhibitors tend to rapidly dissociate from the Top1cc before DNA replication or transcription collision. (B) Collision between a replication fork and a stalled Top1cc (shown on the leading strand) produces a replication-induced double-strand end (“replication run-off”). Similarly, a stalled Top1cc can produce a transcription block and activate the DNA damage response., The repair of covalent Top1-DNA complexes is believed to involve two main pathways. (C) Tdp1 can hydrolyze the tyrosyl-phosphoester bond following the degradation of Top1 by ubiquitin-proteasomal degradation (for review see22,121). (D) Data obtained from yeast DNA repair mutant suggest an alternative endonuclease pathway, which is not yet validated in human cells. The candidate endonucleases are Mre11-CtIP, XPF-ERCC1 and Mus81-Eme1 (for review see2,22).

(A) In normal cells, Top1-DNA covalent complexes can be repaired by redundant mechanisms, which can be divided in two main pathways: i/the Tdp1 hydrolysis pathway (see Figure 5C), and ii/the 3′-endonuclease pathway (see Figure 5D). (B) Cancer cells might be more dependent on the Tdp1 pathway as a result of mutations and inactivation of DNA checkpoints (BRCA1, Chk2…). The expected effect of combining a Tdp1 inhibitor with a Top1 inhibitor would be an increase in the therapeutic index of the Top1 inhibitor as the Tdp1 inhibitor would sensitize preferentially the cancer cells.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
