

Review
doi: 10.1111/j.1742-4658.2009.06985.x.
Epub 2009 Mar 24.
Typical 2-Cys peroxiredoxins--structures, mechanisms and functions
Affiliations
- PMID: 19476488
- PMCID: PMC2747500
- DOI: 10.1111/j.1742-4658.2009.06985.x
Item in Clipboard
Review
Typical 2-Cys peroxiredoxins--structures, mechanisms and functions
FEBS J.
2009 May.
Abstract
Peroxiredoxins are abundant cellular antioxidant proteins that help to control intracellular peroxide levels. These proteins may also function, in part, through an evolved sensitivity of some peroxiredoxins towards peroxide-mediated inactivation in hydrogen peroxide signaling in eukaryotes. This review summarizes recent progress in our understanding of the catalytic and regulatory mechanisms of 'typical 2-Cys' peroxiredoxins and of the biological roles played by these important enzymes in oxidative stress and nonstress-related cellular signaling. New evidence suggests localized peroxide buildup plays a role in nonstress-related signaling.
Figures
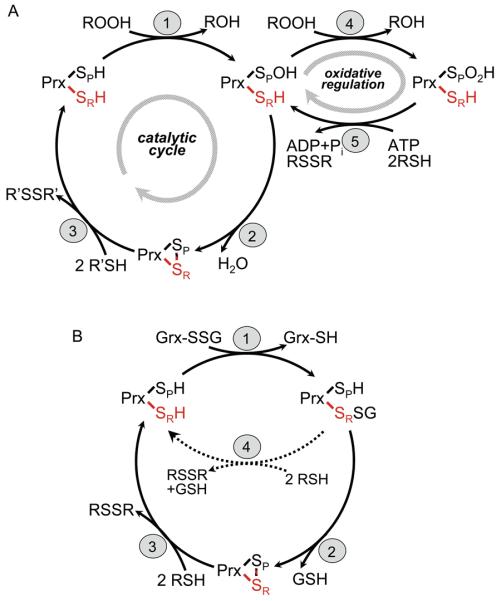
Mechanisms of catalysis by typical 2-Cys Prxs. (A) The peroxidatic catalytic cycle of typical 2-Cys Prxs involves three main steps: (1) peroxidation, (2) resolution and (3) recycling. Not shown is the local unfolding event that occurs in both the CP-loop and C-terminus during step 2 so that the disulfide bond can form (see Figure 2a). The protein is represented as one of two active sites within a functional dimer, with SP and SR (red) designating the sulfur atoms of the peroxidatic and resolving cysteines, respectively, from different subunits. “2 R′SH” in step 3 represents a thioredoxin-like protein or domain. Overoxidation of the CP (step 4) and reduction of the Cys-SPO2H by Srx (step 5) depict redox regulation and repair occurring in some eukaryotic typical 2-Cys Prxs. (B) Mutants of AhpC which suppress the growth defect (dithiothreitol dependence) of the trxB gor mutant from E. coli were shown to catalyze the deglutathionylation of Grx1 (using the C14S mutant) in vitro [24]. Although the catalytic intermediate is shown with glutathione attached in a mixed disulfide to the Prx, the alternative mechanism with Grx attached to Prx is also possible. The truncated cycle shown with dotted lines (step 4 in b) illustrates the finding that only the “resolving cysteine” (with the sulfur depicted as SR) is required for this activity.
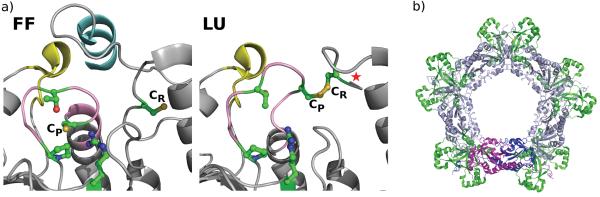
Structural aspects of Prx sensitivity and resurrection. (A) The active site of a sensitive 2-Cys Prx in the fully folded (FF) and locally unfolded (LU) conformations. Both chains of the dimer are colored grey. The C-terminal helix containing the YF motif (cyan) and the loop associated with the GGLG motif (yellow) that are characteristic of sensitive Prxs can be seen to pack against each other and cover the active site CP loop (pink) in the FF conformation (PDB code 1QMV). Comparing the LU structure (PDB code 1QQ2) with the FF structure shows how the CP loop and the protein C-terminus unfold for disulfide formation and how the C-terminal helix hinders this required local unfolding. In robust Prxs, the C-terminal helix containing the YF motif is absent, allowing for more facile unfolding (see Figure 2 in Wood et al, 2003). In the LU form, a star indicates the presence of the additional disordered C-terminal residues. In both the FF and LU images, the four residues conserved in all Prxs (CP, Arg, Thr and Pro) and the CR are colored green with sulfur, oxygen and nitrogen atoms colored dark yellow, red and blue, respectively. (B) The typical 2-Cys dimer (magenta and dark blue) associates with other dimers (light blue) as part of the normal catalytic cycle to form higher order oligomers. The overoxidized state is stabilized in this form [20]. Modeling of Srxs (green) on a Prx decamer shows that Srx can associate with such a structure without significant changes to the decamer, consistent with the role of Srx in the reduction of the overoxidized Prx. Figure modified from [32].
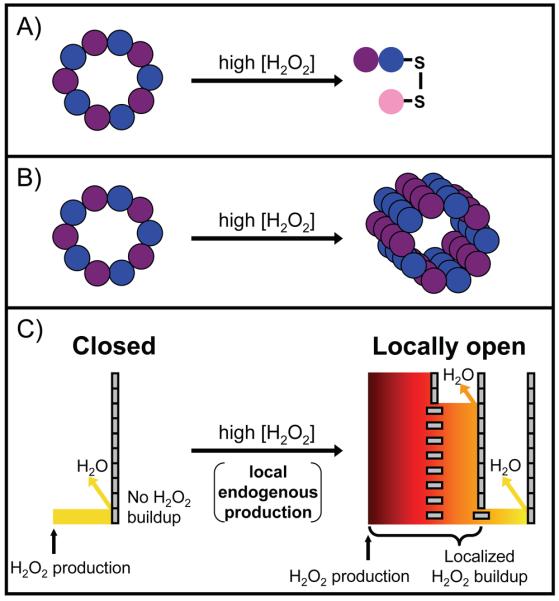
Three proposed roles for Prxs in peroxide signaling. In each case, different high levels of hydrogen peroxide cause a shift in function from peroxidase activity. (A) Disulfide exchange, represented by an interprotein disulfide bond between Prx and a downstream protein (pink). In the one case studied, signaling is not stress related and does not require sensitivity. (B) The chaperone model, represented by the formation of higher order oligomers of overoxidized Prxs. This is involved in stress related signaling and requires sensitivity. In (A) and (B), the Prxs are represented as a purple and blue decamer under normal cellular conditions. (C) The floodgate model is an unproven mechanism. The Prxs are represented as tall barriers made up of gray rectangles – vertical for active, horizontal for overoxidized and inactive. The multiple barriers on the right reflect the cell-wide Prx distribution; Prxs that are close to the peroxide generation site (marked by an arrow) are overwhelmed and inactivated, whereas those at increasing distances away are not. This creates a steep peroxide gradient and allows for localized peroxide buildup after endogenous peroxide generation. The level of hydrogen peroxide is represented by both color gradient and height. This may be involved in both stress and non-stress related signaling and requires sensitivity.
References
-
- Wood ZA, Schröder E, Harris JR, Poole LB. Structure, mechanism and regulation of peroxiredoxins. Trends Biochem Sci. 2003;28:32–40. - PubMed
-
- Copley SD, Novak WR, Babbitt PC. Divergence of function in the thioredoxin fold suprafamily: evidence for evolution of peroxiredoxins from a thioredoxin-like ancestor. Biochemistry. 2004;43:13981–13995. - PubMed
-
- Chae HZ, Chung SJ, Rhee SG. Thioredoxin-dependent peroxide reductase from yeast. J Biol Chem. 1994;269:27670–27678. - PubMed
-
- Fourquet S, Huang ME, D'Autreaux B, Toledano MB. The dual functions of thiol-based peroxidases in H2O2 scavenging and signaling. Antioxid Redox Signal. 2008;10:1565–1576. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
