

Biases in Drosophila melanogaster protein trap screens
- PMID: 19476619
- PMCID: PMC2695487
- DOI: 10.1186/1471-2164-10-249
Biases in Drosophila melanogaster protein trap screens
Abstract
Background: The ability to localise or follow endogenous proteins in real time in vivo is of tremendous utility for cell biology or systems biology studies. Protein trap screens utilise the random genomic insertion of a transposon-borne artificial reporter exon (e.g. encoding the green fluorescent protein, GFP) into an intron of an endogenous gene to generate a fluorescent fusion protein. Despite recent efforts aimed at achieving comprehensive coverage of the genes encoded in the Drosophila genome, the repertoire of genes that yield protein traps is still small.
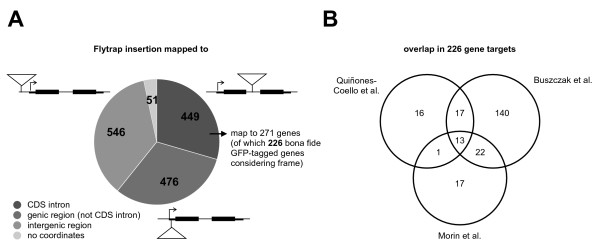
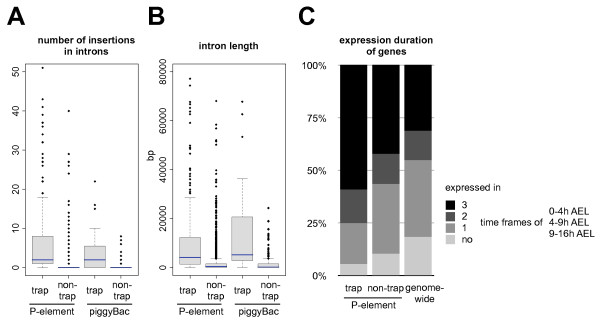
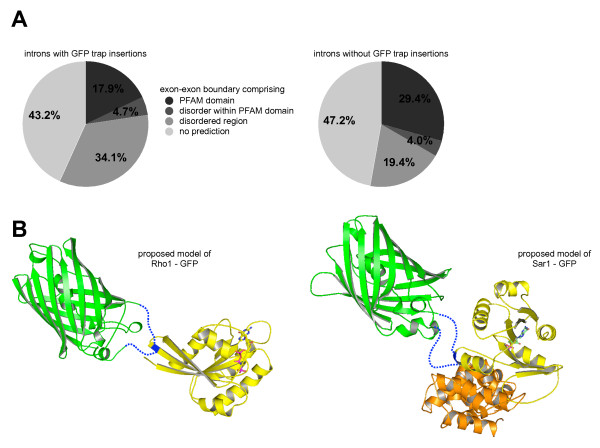
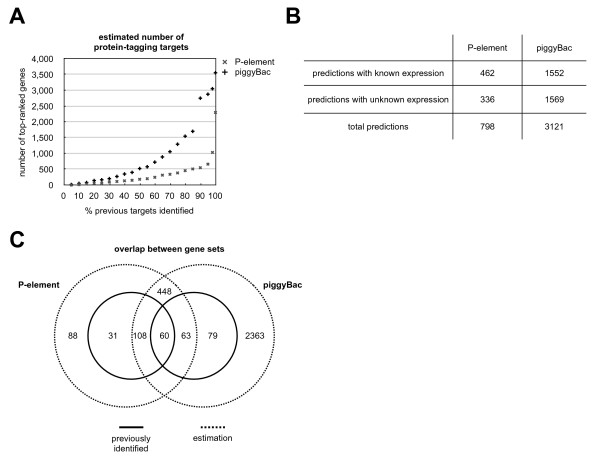
Results: We analysed the collection of available protein trap lines in Drosophila melanogaster and identified potential biases that are likely to restrict genome coverage in protein trap screens. The protein trap screens investigated here primarily used P-element vectors and thus exhibit some of the same positional biases associated with this transposon that are evident from the comprehensive Drosophila Gene Disruption Project. We further found that protein trap target genes usually exhibit broad and persistent expression during embryonic development, which is likely to facilitate better detection. In addition, we investigated the likely influence of the GFP exon on host protein structure and found that protein trap insertions have a significant bias for exon-exon boundaries that encode disordered protein regions. 38.8% of GFP insertions land in disordered protein regions compared with only 23.4% in the case of non-trapping P-element insertions landing in coding sequence introns (p < 10(-4)). Interestingly, even in cases where protein domains are predicted, protein trap insertions frequently occur in regions encoding surface exposed areas that are likely to be functionally neutral. Considering the various biases observed, we predict that less than one third of intron-containing genes are likely to be amenable to trapping by the existing methods.
Conclusion: Our analyses suggest that the utility of P-element vectors for protein trap screens has largely been exhausted, and that approximately 2,800 genes may still be amenable using piggyBac vectors. Thus protein trap strategies based on current approaches are unlikely to offer true genome-wide coverage. We suggest that either transposons with reduced insertion bias or recombineering-based targeting techniques will be required for comprehensive genome coverage in Drosophila.
Figures
Similar articles
-
Exploring strategies for protein trapping in Drosophila.Genetics. 2007 Mar;175(3):1089-104. doi: 10.1534/genetics.106.065995. Epub 2006 Dec 18. Genetics. 2007. PMID: 17179094 Free PMC article.
-
Minos as a genetic and genomic tool in Drosophila melanogaster.Genetics. 2005 Oct;171(2):571-81. doi: 10.1534/genetics.105.041848. Epub 2005 Jun 21. Genetics. 2005. PMID: 15972463 Free PMC article.
-
The carnegie protein trap library: a versatile tool for Drosophila developmental studies.Genetics. 2007 Mar;175(3):1505-31. doi: 10.1534/genetics.106.065961. Epub 2006 Dec 28. Genetics. 2007. PMID: 17194782 Free PMC article.
-
Transposable elements as tools for genomics and genetics in Drosophila.Brief Funct Genomic Proteomic. 2003 Apr;2(1):57-71. doi: 10.1093/bfgp/2.1.57. Brief Funct Genomic Proteomic. 2003. PMID: 15239944 Review.
-
deGradFP: A System to Knockdown GFP-Tagged Proteins.Methods Mol Biol. 2016;1478:177-187. doi: 10.1007/978-1-4939-6371-3_9. Methods Mol Biol. 2016. PMID: 27730581 Review.
Cited by
-
A library of MiMICs allows tagging of genes and reversible, spatial and temporal knockdown of proteins in Drosophila.Elife. 2015 Mar 31;4:e05338. doi: 10.7554/eLife.05338. Elife. 2015. PMID: 25824290 Free PMC article.
-
Inducible protein traps with dominant phenotypes for functional analysis of the Drosophila genome.Genetics. 2014 Jan;196(1):91-105. doi: 10.1534/genetics.113.157529. Epub 2013 Oct 30. Genetics. 2014. PMID: 24172131 Free PMC article.
-
In vivo analysis of proteomes and interactomes using Parallel Affinity Capture (iPAC) coupled to mass spectrometry.Mol Cell Proteomics. 2011 Jun;10(6):M110.002386. doi: 10.1074/mcp.M110.002386. Epub 2011 Mar 29. Mol Cell Proteomics. 2011. PMID: 21447707 Free PMC article.
-
In vivo protein trapping produces a functional expression codex of the vertebrate proteome.Nat Methods. 2011 Jun;8(6):506-15. doi: 10.1038/nmeth.1606. Epub 2011 May 8. Nat Methods. 2011. PMID: 21552255 Free PMC article.
-
Stringent analysis of gene function and protein-protein interactions using fluorescently tagged genes.Genetics. 2012 Mar;190(3):931-40. doi: 10.1534/genetics.111.136465. Epub 2011 Dec 14. Genetics. 2012. PMID: 22174071 Free PMC article.
References
-
- Thibault ST, Singer MA, Miyazaki WY, Milash B, Dompe NA, Singh CM, Buchholz R, Demsky M, Fawcett R, Francis-Lang HL, Ryner L, Cheung LM, Chong A, Erickson C, Fisher WW, Greer K, Hartouni SR, Howie E, Jakkula L, Joo D, Killpack K, Laufer A, Mazzotta J, Smith RD, Stevens LM, Stuber C, Tan LR, Ventura R, Woo A, Zakrajsek I, Zhao L, Chen F, Swimmer C, Kopczynski C, Duyk G, Winberg ML, Margolis J. A complementary transposon tool kit for Drosophila melanogaster using P and piggyBac. Nat Genet. 2004;36:283–7. doi: 10.1038/ng1314. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
