

A general model of synaptic transmission and short-term plasticity
- PMID: 19477155
- PMCID: PMC3035647
- DOI: 10.1016/j.neuron.2009.03.025
A general model of synaptic transmission and short-term plasticity
Abstract
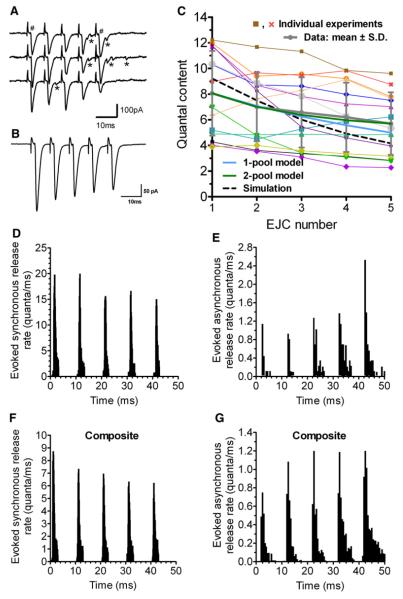
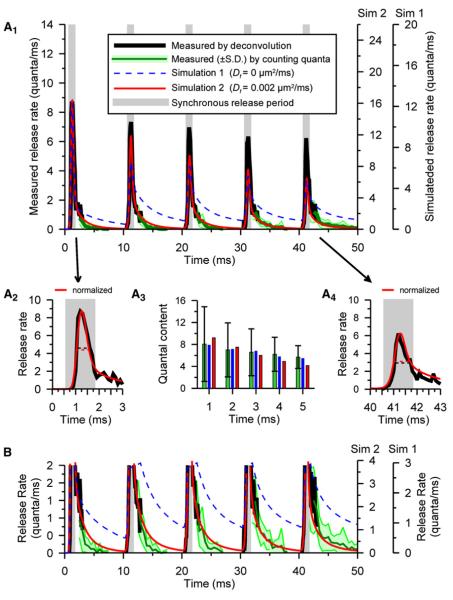
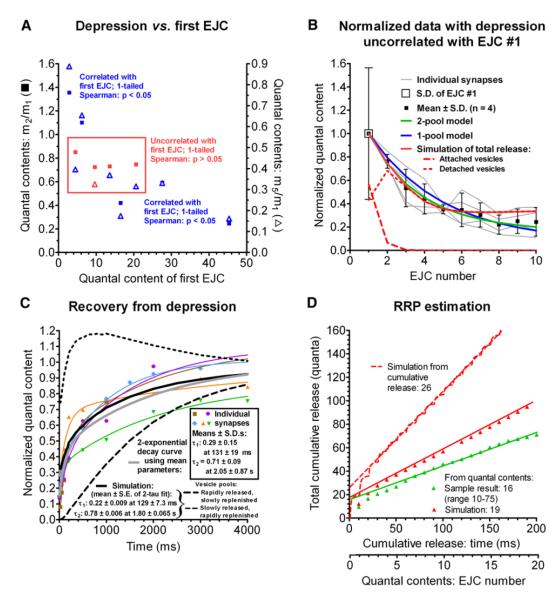
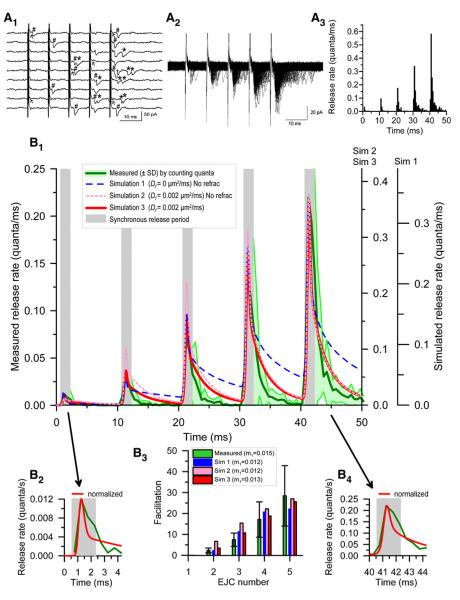
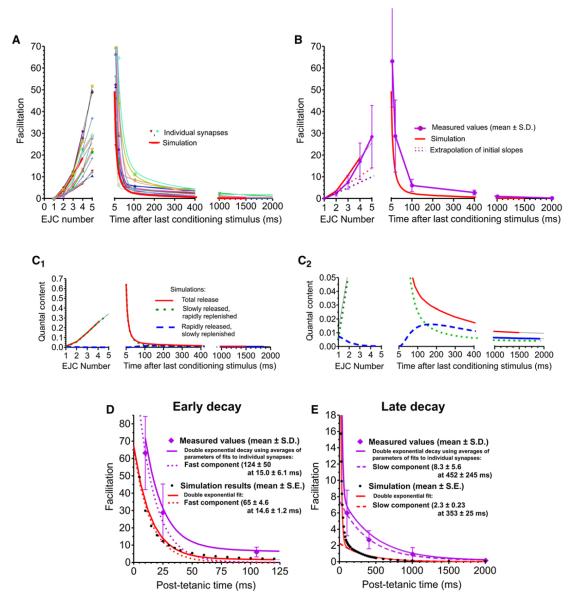
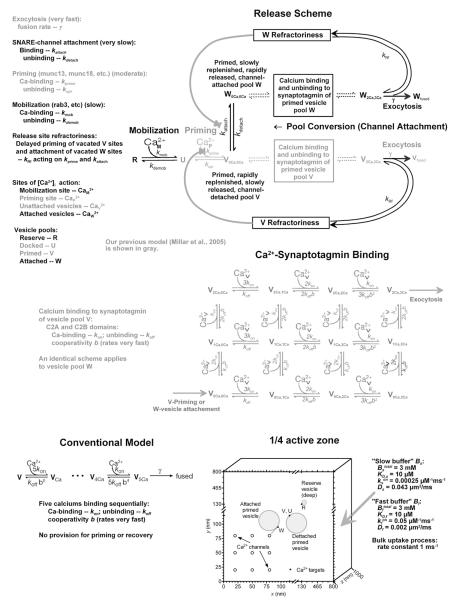
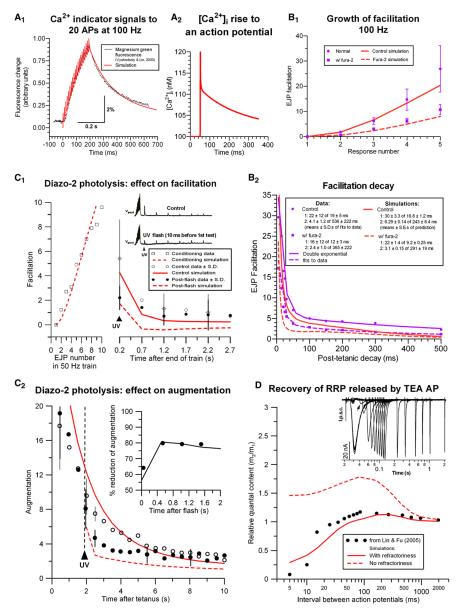
Some synapses transmit strongly to action potentials (APs), but weaken with repeated activation; others transmit feebly at first, but strengthen with sustained activity. We measured synchronous and asynchronous transmitter release at "phasic" crayfish neuromuscular junctions (NMJs) showing depression and at facilitating "tonic" junctions, and define the kinetics of depression and facilitation. We offer a comprehensive model of presynaptic processes, encompassing mobilization of reserve vesicles, priming of docked vesicles, their association with Ca(2+) channels, and refractoriness of release sites, while accounting for data on presynaptic buffers governing Ca(2+) diffusion. Model simulations reproduce many experimentally defined aspects of transmission and plasticity at these synapses. Their similarity to vertebrate central synapses suggests that the model might be of general relevance to synaptic transmission.
Figures
References
-
- Adams SR, Kao JPY, Tsien RY. Biologically useful chelators that take up Ca2+ upon illumination. J. Am. Chem. Soc. 1989;111:7957–7968.
-
- Atwood HL, Bittner GD. Matching of excitatory and inhibitory inputs to crustacean muscle fibers. J. Neurophysiol. 1971;34:157–170. - PubMed
-
- Atwood HL, Karunanithi S. Diversification of synaptic strength: presynaptic elements. Nat. Rev. Neurosci. 2002;3:497–516. - PubMed
-
- Beaumont V, Zhong N, Froemke RC, Ball RW, Zucker RS. Temporal synaptic tagging by Ih activation and actin: involvement in long-term facilitation and cAMP-induced synaptic enhancement. Neuron. 2002;33:601–613. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
