

Conservation of proteins involved in oocyst wall formation in Eimeria maxima, Eimeria tenella and Eimeria acervulina
- PMID: 19477178
- PMCID: PMC2726925
- DOI: 10.1016/j.ijpara.2009.05.004
Conservation of proteins involved in oocyst wall formation in Eimeria maxima, Eimeria tenella and Eimeria acervulina
Abstract
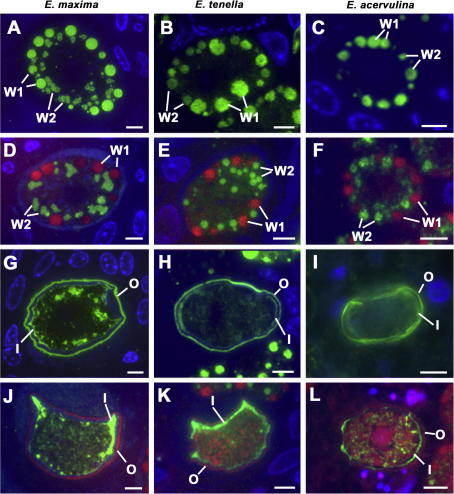
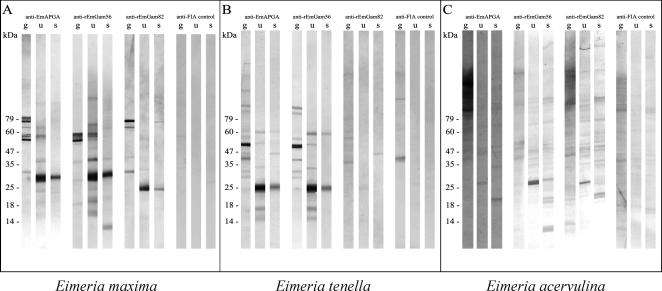
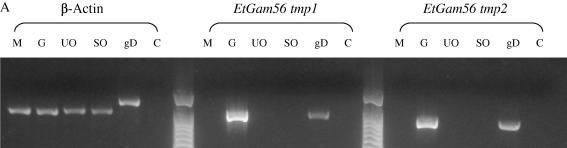
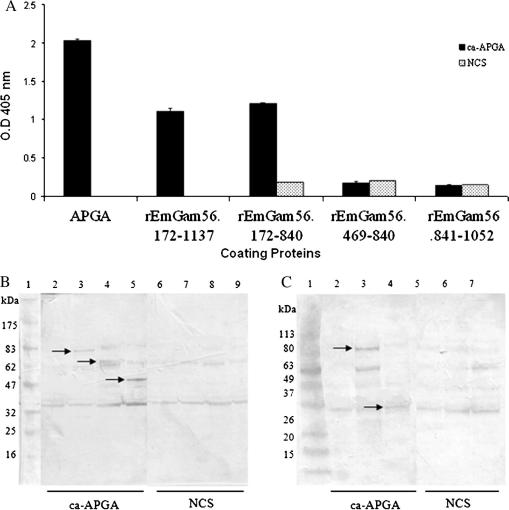
Vaccination with proteins from gametocytes of Eimeria maxima protects chickens, via transfer of maternal antibodies, against infection with several species of Eimeria. Antibodies to E. maxima gametocyte proteins recognise proteins in the wall forming bodies of macrogametocytes and oocyst walls of E. maxima, Eimeria tenella and Eimeria acervulina. Homologous genes for two major gametocyte proteins - GAM56 and GAM82 - were found in E. maxima, E. tenella and E. acervulina. Alignment of the predicted protein sequences of these genes reveals that, as well as sharing regions of tyrosine richness, strong homology exists in their amino-terminal regions, where protective antibodies bind. This study confirms the conservation of the roles of GAM56 and GAM82 in oocyst wall formation and shows that antibodies to gametocyte antigens of E. maxima cross-react with homologous proteins in other species, helping to explain cross-species maternal immunity.
Figures

References
-
- Belli S.I., Lee M., Thebo P., Wallach M.G., Schwartsburd B., Smith N.C. Biochemical characterisation of the 56 and 82 kDa immunodominant gametocyte antigens from Eimeria maxima. Int. J. Parasitol. 2002;32:805–816. - PubMed
-
- Belli S.I., Witcombe D., Wallach M.G., Smith N.C. Functional genomics of gam56: characterisation of the role of a 56 kDa sexual stage antigen in oocyst wall formation in Eimeria maxima. Int. J. Parasitol. 2002;32:1727–1737. - PubMed
-
- Belli S.I., Wallach M.G., Luxford C., Davies M.J., Smith N.C. Roles of tyrosine-rich precursor glycoproteins and dityrosine- and 3,4-dihydroxyphenylalanine-mediated protein cross-linking in development of the oocyst wall in the coccidian parasite Eimeria maxima. Eukaryot. Cell. 2003;2:456–464. - PMC - PubMed
-
- Belli S.I., Wallach M.G., Smith N.C. Cloning and characterization of the 82 kDa tyrosine-rich sexual stage glycoprotein, GAM82, and its role in oocyst wall formation in the apicomplexan parasite, Eimeria maxima. Gene. 2003;307:201–212. - PubMed
-
- Belli S.I., Mai K., Skene C.D., Gleeson M.T., Witcombe D.M., Katrib M., Finger A., Wallach M.G., Smith N.C. Characterisation of the antigenic and immunogenic properties of bacterially expressed, sexual stage antigens of the coccidian parasite, Eimeria maxima. Vaccine. 2004;22:4316–4325. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
