

CD152 (CTLA-4) determines CD4 T cell migration in vitro and in vivo
- PMID: 19479036
- PMCID: PMC2682661
- DOI: 10.1371/journal.pone.0005702
CD152 (CTLA-4) determines CD4 T cell migration in vitro and in vivo
Abstract
Background: Migration of antigen-experienced T cells to secondary lymphoid organs and the site of antigenic-challenge is a mandatory prerequisite for the precise functioning of adaptive immune responses. The surface molecule CD152 (CTLA-4) is mostly considered as a negative regulator of T cell activation during immune responses. It is currently unknown whether CD152 can also influence chemokine-driven T cell migration.
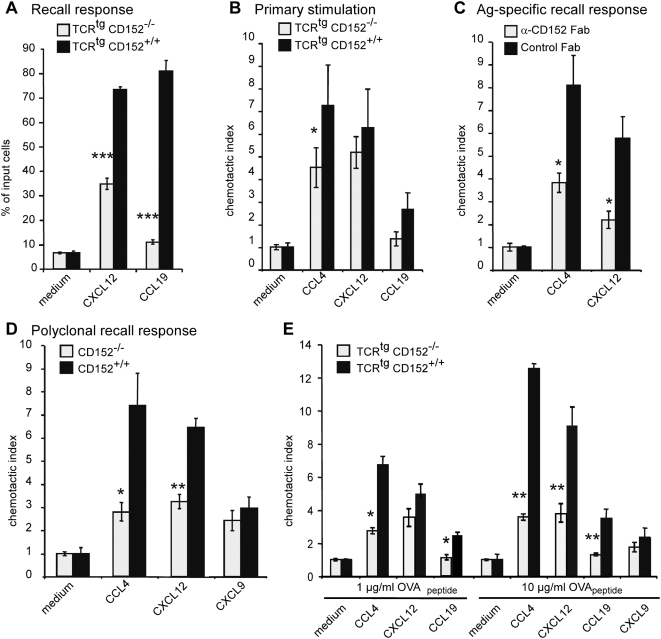
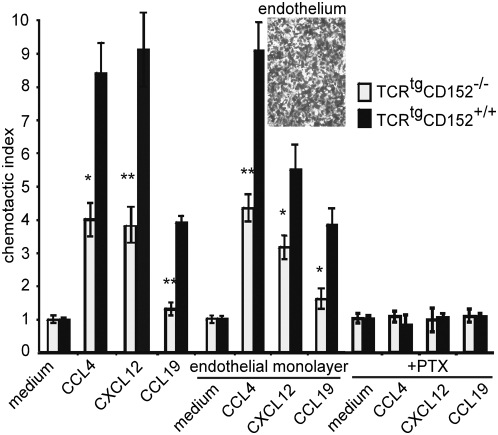
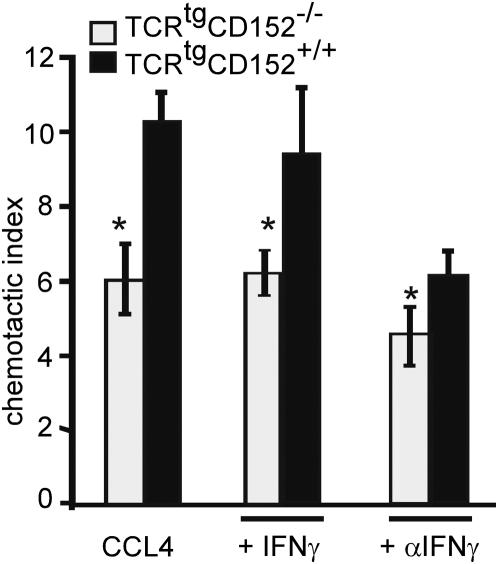
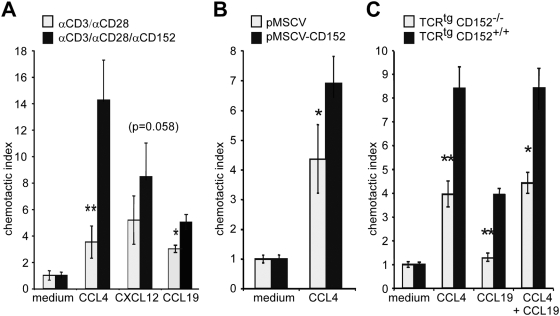
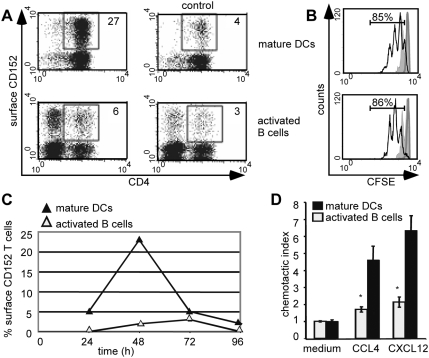
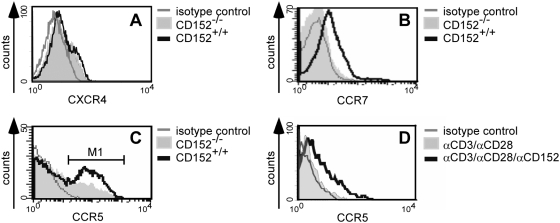
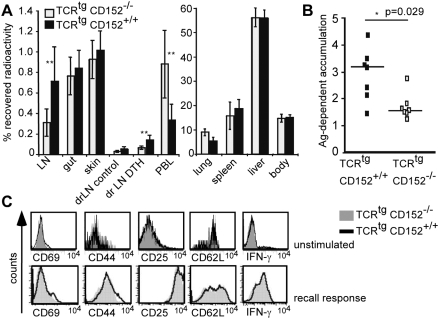
Methodology/principal findings: We analyzed the consequences of CD152 signaling on Th cell migration using chemotaxis assays in vitro and radioactive cell tracking in vivo. We show here that the genetic and serological inactivation of CD152 in Th1 cells reduced migration towards CCL4, CXCL12 and CCL19, but not CXCL9, in a G-protein dependent manner. In addition, retroviral transduction of CD152 cDNA into CD152 negative cells restored Th1 cell migration. Crosslinking of CD152 together with CD3 and CD28 stimulation on activated Th1 cells increased expression of the chemokine receptors CCR5 and CCR7, which in turn enhanced cell migration. Using sensitive liposome technology, we show that mature dendritic cells but not activated B cells were potent at inducing surface CD152 expression and the CD152-mediated migration-enhancing signals. Importantly, migration of CD152 positive Th1 lymphocytes in in vivo experiments increased more than 200% as compared to CD152 negative counterparts showing that indeed CD152 orchestrates specific migration of selected Th1 cells to sites of inflammation and antigenic challenge in vivo.
Conclusions/significance: We show here, that CD152 signaling does not just silence cells, but selects individual ones for migration. This novel activity of CD152 adds to the already significant role of CD152 in controlling peripheral immune responses by allowing T cells to localize correctly during infection. It also suggests that interference with CD152 signaling provides a tool for altering the cellular composition at sites of inflammation and antigenic challenge.
Conflict of interest statement
Figures
References
-
- Campbell DJ, Kim CH, Butcher EC. Chemokines in the systemic organization of immunity. Immunological Reviews. 2003;195:58–71. - PubMed
-
- Hamann A, Engelhardt B. Leukocyte trafficking. Weinheim: Wiley-VCH; 2005. p. 517.
-
- Szabo SJ, Sullivan BM, Peng SL, Glimcher LH. Molecular mechanisms regulating Th1 immune responses. Annual Review of Immunology. 2003;21:713–758. - PubMed
-
- Olsson T. Critical Influences of the Cytokine Orchestration on the Outcome of Myelin Antigen-Specific T-Cell Autoimmunity in Experimental Autoimmune Encephalomyelitis and Multiple-Sclerosis. Immunological Reviews. 1995;144:245–268. - PubMed
-
- Casares D, Brumeanu TD. Insights into the pathogeneses of type I diabetes: a hint for novel immunospecific therapies. Curr Mol Med. 2001;1:357–378. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
