

In vivo reorganization of the actin cytoskeleton in leaves of Nicotiana tabacum L. transformed with plastin-GFP. Correlation with light-activated chloroplast responses
- PMID: 19480655
- PMCID: PMC2702303
- DOI: 10.1186/1471-2229-9-64
In vivo reorganization of the actin cytoskeleton in leaves of Nicotiana tabacum L. transformed with plastin-GFP. Correlation with light-activated chloroplast responses
Abstract
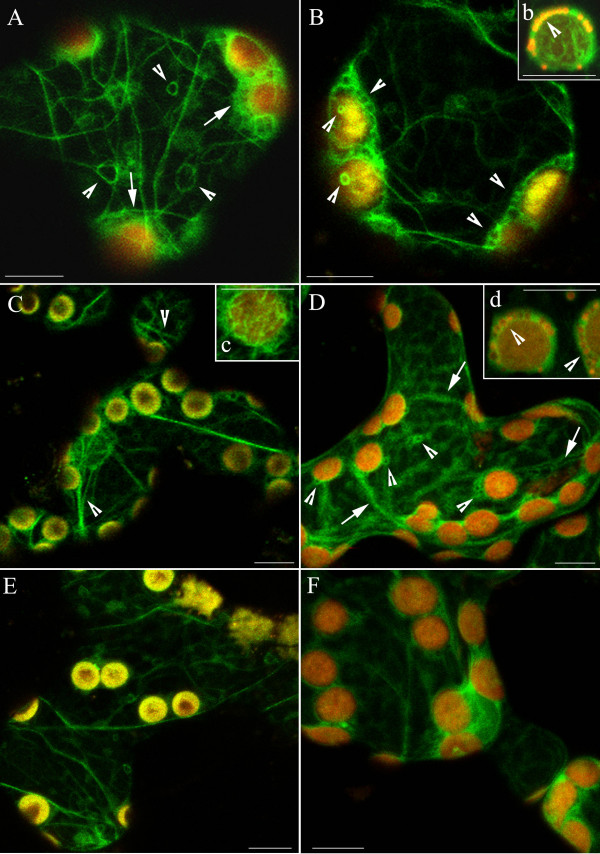
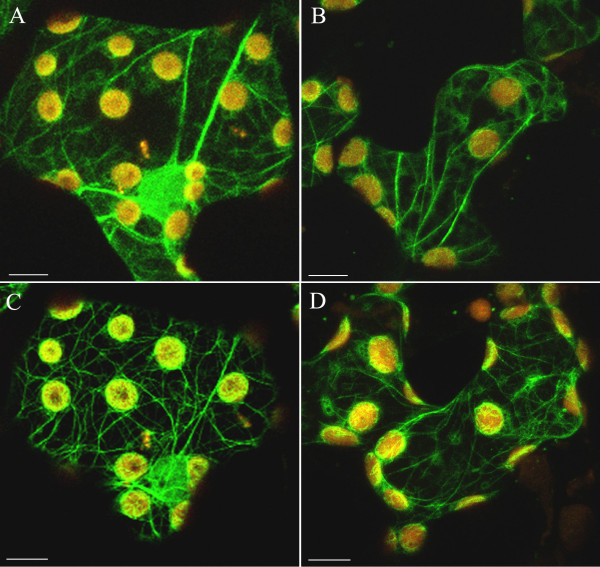
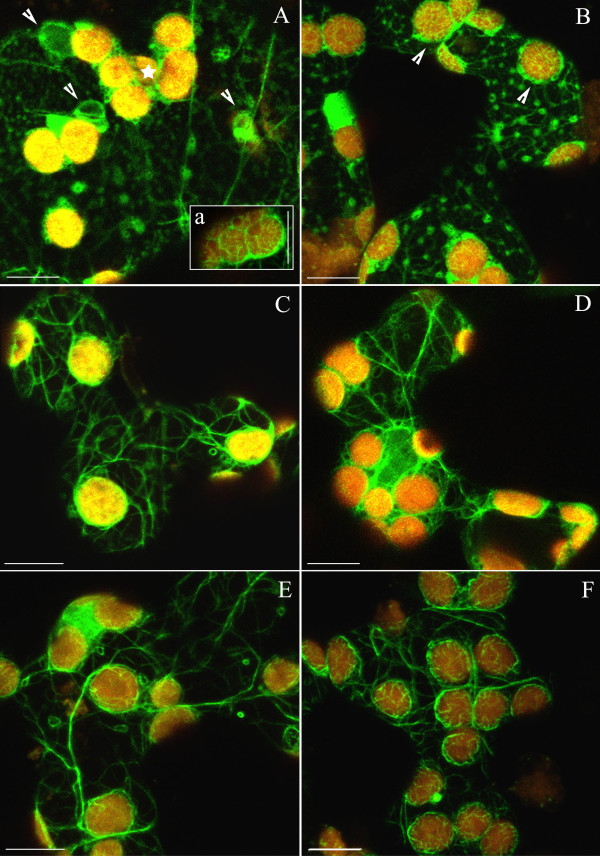
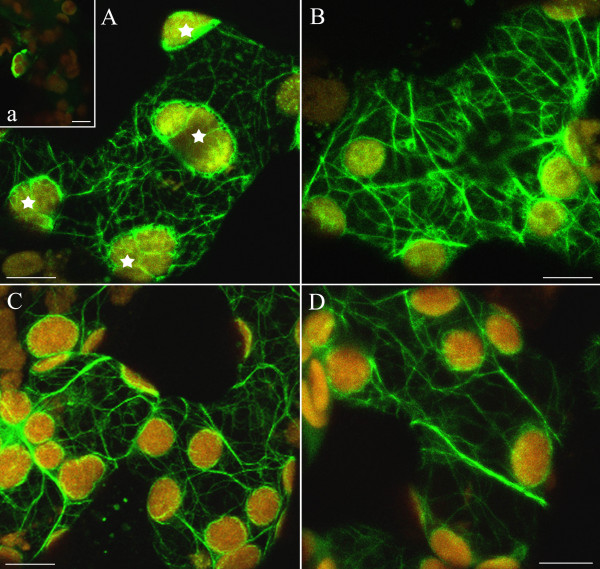
Background: The actin cytoskeleton is involved in the responses of plants to environmental signals. Actin bundles play the role of tracks in chloroplast movements activated by light. Chloroplasts redistribute in response to blue light in the mesophyll cells of Nicotiana tabacum. The aim of this work was to study the relationship between chloroplast responses and the organization of actin cytoskeleton in living tobacco cells. Chloroplast movements were measured photometrically as changes in light transmission through the leaves. The actin cytoskeleton, labeled with plastin-GFP, was visualised by confocal microscopy.
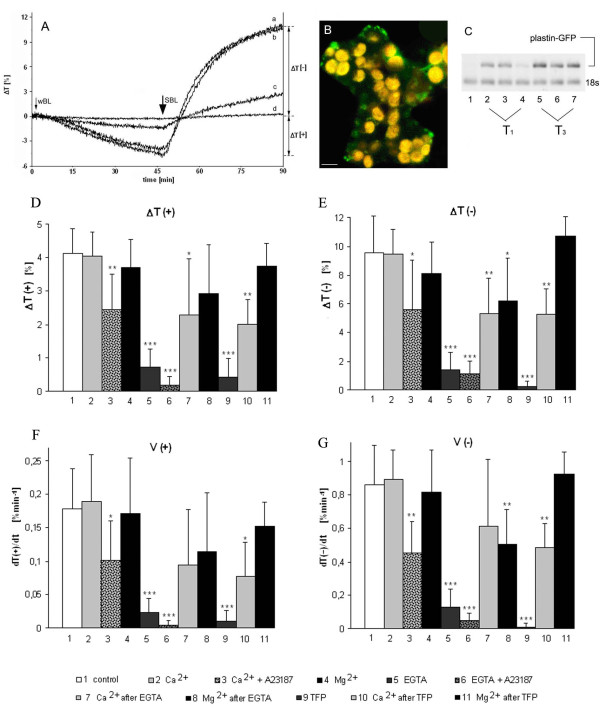
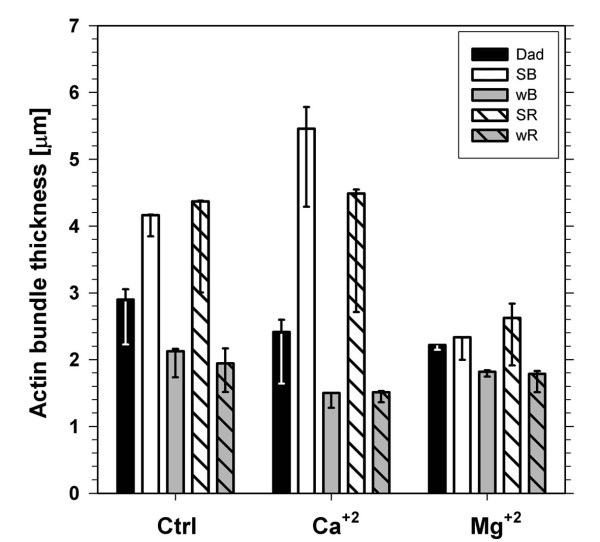
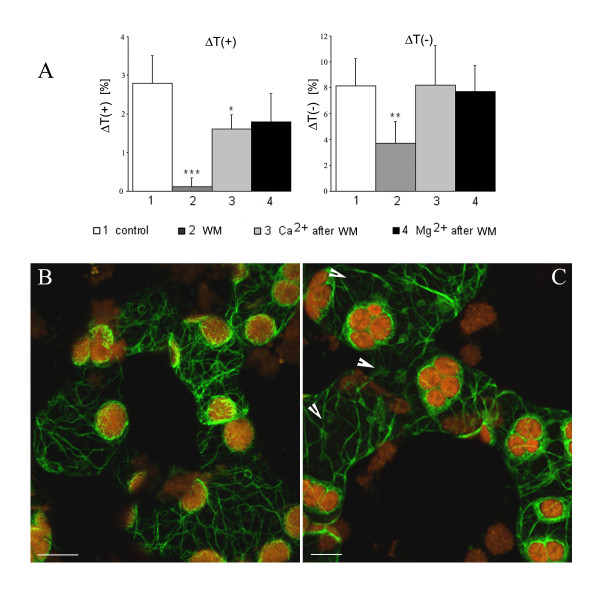
Results: The actin cytoskeleton was affected by strong blue and red light. No blue light specific actin reorganization was detected. EGTA and trifluoperazine strongly inhibited chloroplast responses and disrupted the integrity of the cytoskeleton. This disruption was reversible by Ca(2+) or Mg(2+). Additionally, the effect of trifluoperazine was reversible by light. Wortmannin, an inhibitor of phosphoinositide kinases, potently inhibited chloroplast responses but did not influence the actin cytoskeleton at the same concentration. Also this inhibition was reversed by Ca(2+) and Mg(2+). Magnesium ions were equally or more effective than Ca(2+) in restoring chloroplast motility after treatment with EGTA, trifluoperazine or wortmannin.
Conclusion: The architecture of the actin cytoskeleton in the mesophyll of tobacco is significantly modulated by strong light. This modulation does not affect the direction of chloroplast redistribution in the cell. Calcium ions have multiple functions in the mechanism of the movements. Our results suggest also that Mg(2+) is a regulatory molecule cooperating with Ca(2+) in the signaling pathway of blue light-induced tobacco chloroplast movements.
Figures
References
-
- Grolig F. Organelle movements: transport and positioning. In: Hussey PJ, editor. The plant cytoskeleton in cell differentiation and development. Oxford, Blackwell Publishing; 2004. pp. 148–175.
-
- Haupt W, Scheuerlein R. Chloroplast movement. Plant Cell Environ. 1990;13:596–614. doi: 10.1111/j.1365-3040.1990.tb01078.x. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
