

Influenza virus morphogenesis and budding
- PMID: 19481124
- PMCID: PMC2730999
- DOI: 10.1016/j.virusres.2009.05.010
Influenza virus morphogenesis and budding
Abstract
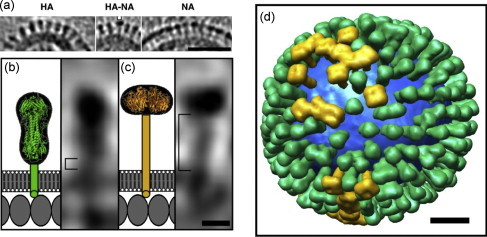
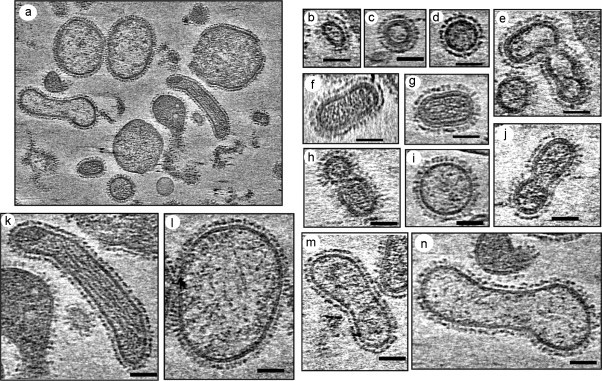
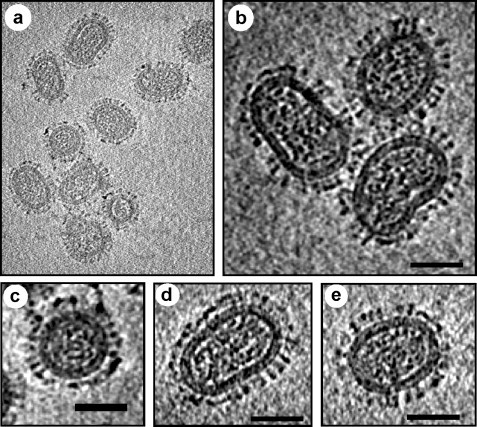
Influenza viruses are enveloped, negative stranded, segmented RNA viruses belonging to Orthomyxoviridae family. Each virion consists of three major sub-viral components, namely (i) a viral envelope decorated with three transmembrane proteins hemagglutinin (HA), neuraminidase (NA) and M2, (ii) an intermediate layer of matrix protein (M1), and (iii) an innermost helical viral ribonucleocapsid [vRNP] core formed by nucleoprotein (NP) and negative strand viral RNA (vRNA). Since complete virus particles are not found inside the cell, the processes of assembly, morphogenesis, budding and release of progeny virus particles at the plasma membrane of the infected cells are critically important for the production of infectious virions and pathogenesis of influenza viruses as well. Morphogenesis and budding require that all virus components must be brought to the budding site which is the apical plasma membrane in polarized epithelial cells whether in vitro cultured cells or in vivo infected animals. HA and NA forming the outer spikes on the viral envelope possess apical sorting signals and use exocytic pathways and lipid rafts for cell surface transport and apical sorting. NP also has apical determinant(s) and is probably transported to the apical budding site similarly via lipid rafts and/or through cortical actin microfilaments. M1 binds the NP and the exposed RNAs of vRNPs, as well as to the cytoplasmic tails (CT) and transmembrane (TM) domains of HA, NA and M2, and is likely brought to the budding site on the piggy-back of vRNP and transmembrane proteins. Budding processes involve bud initiation, bud growth and bud release. The presence of lipid rafts and assembly of viral components at the budding site can cause asymmetry of lipid bilayers and outward membrane bending leading to bud initiation and bud growth. Bud release requires fusion of the apposing viral and cellular membranes and scission of the virus buds from the infected cellular membrane. The processes involved in bud initiation, bud growth and bud scission/release require involvement both viral and host components and can affect bud closing and virus release in both positive and negative ways. Among the viral components, M1, M2 and NA play important roles in bud release and M1, M2 and NA mutations all affect the morphology of buds and released viruses. Disassembly of host cortical actin microfilaments at the pinching-off site appears to facilitate bud fission and release. Bud scission is energy dependent and only a small fraction of virus buds present on the cell surface is released. Discontinuity of M1 layer underneath the lipid bilayer, absence of outer membrane spikes, absence of lipid rafts in the lipid bilayer, as well as possible presence of M2 and disassembly of cortical actin microfilaments at the pinching-off site appear to facilitate bud fission and bud release. We provide our current understanding of these important processes leading to the production of infectious influenza virus particles.
Figures
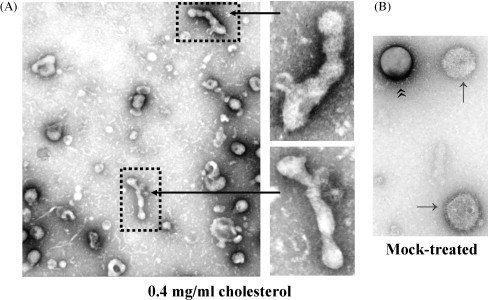
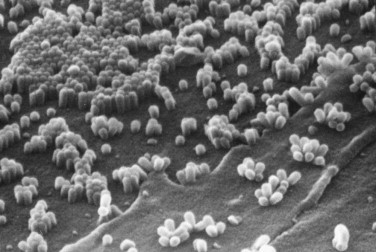

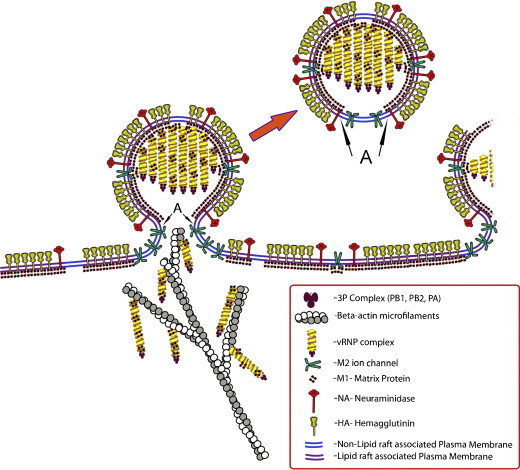
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
