

FACT and Asf1 regulate nucleosome dynamics and coactivator binding at the HO promoter
- PMID: 19481521
- PMCID: PMC2767235
- DOI: 10.1016/j.molcel.2009.04.010
FACT and Asf1 regulate nucleosome dynamics and coactivator binding at the HO promoter
Abstract
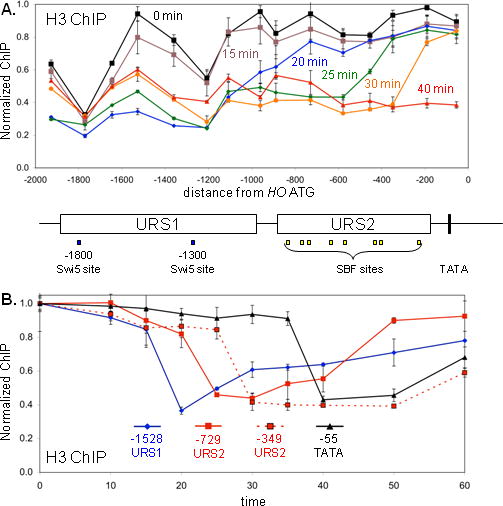
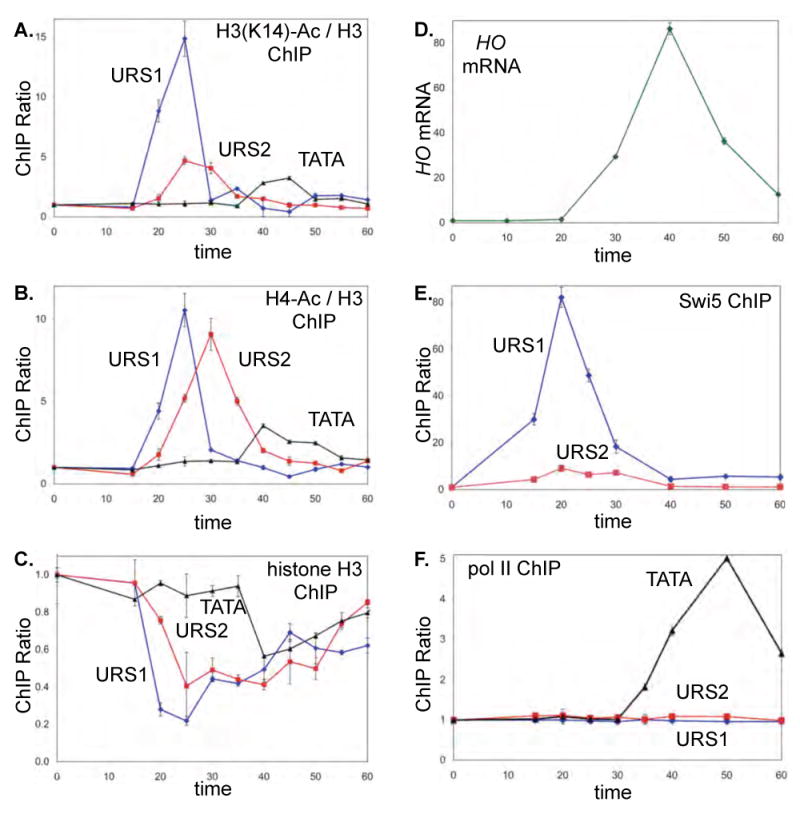
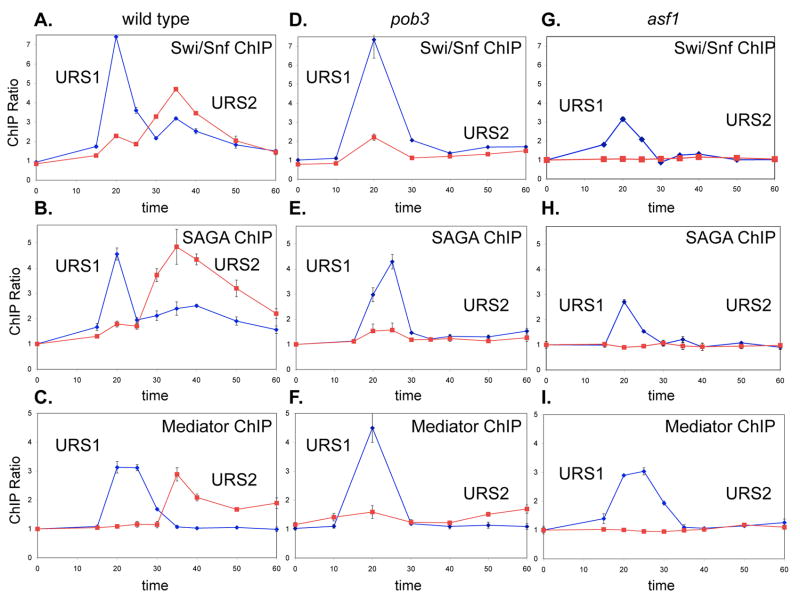
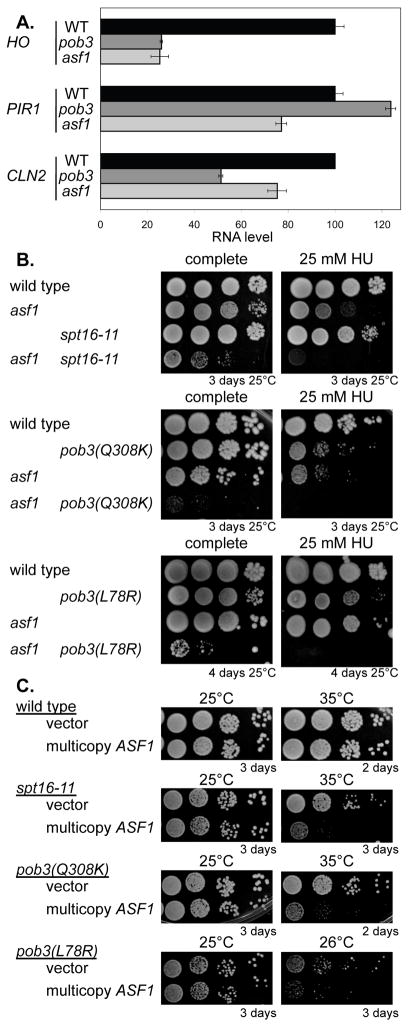
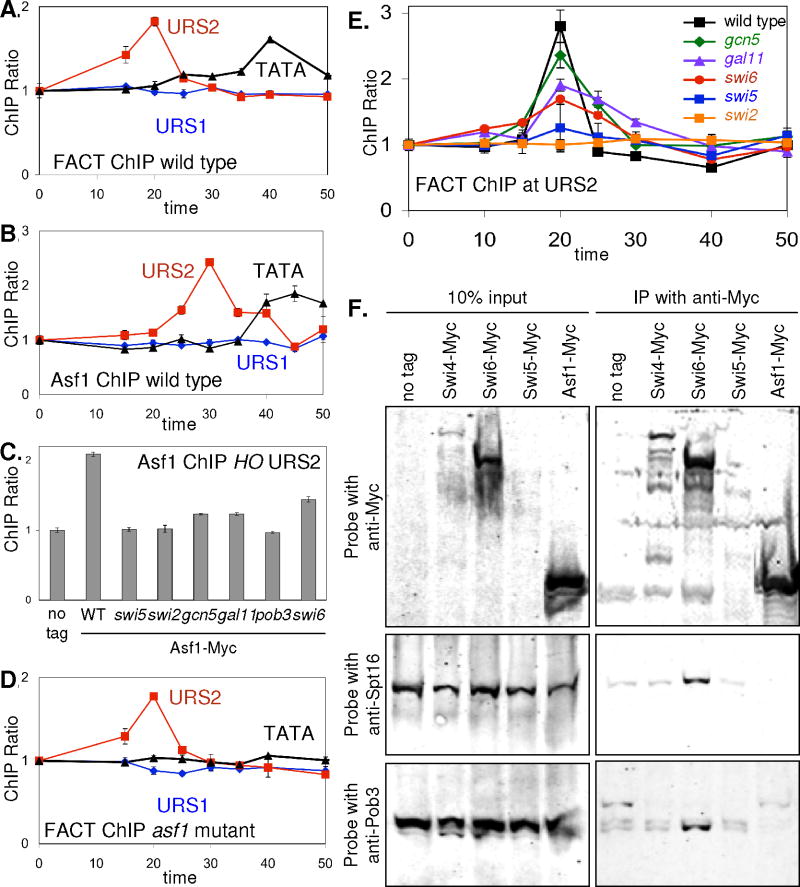
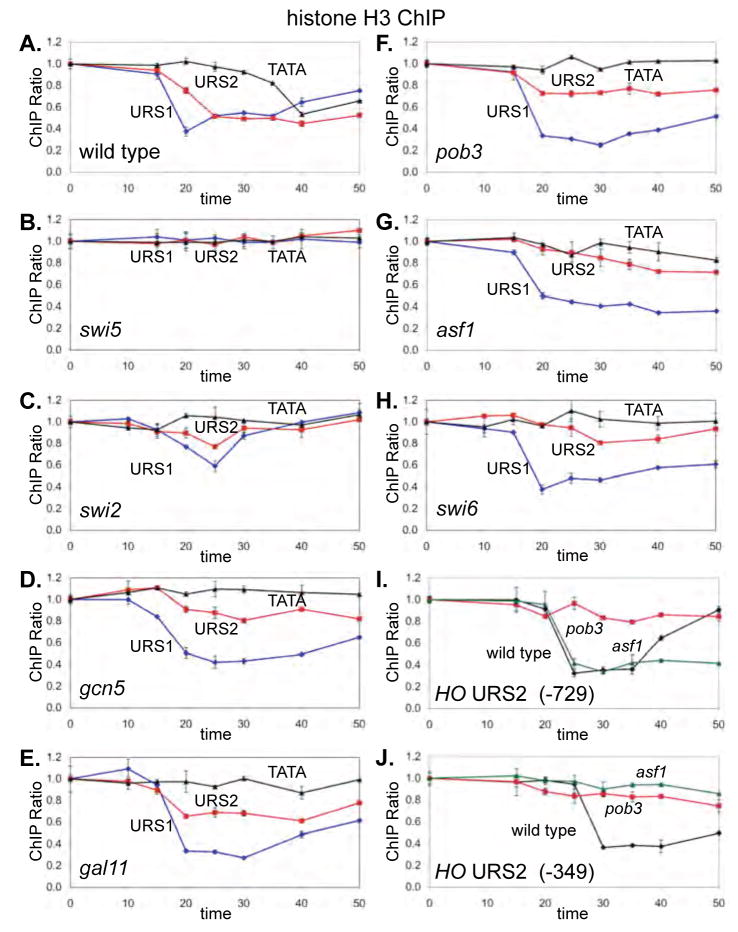
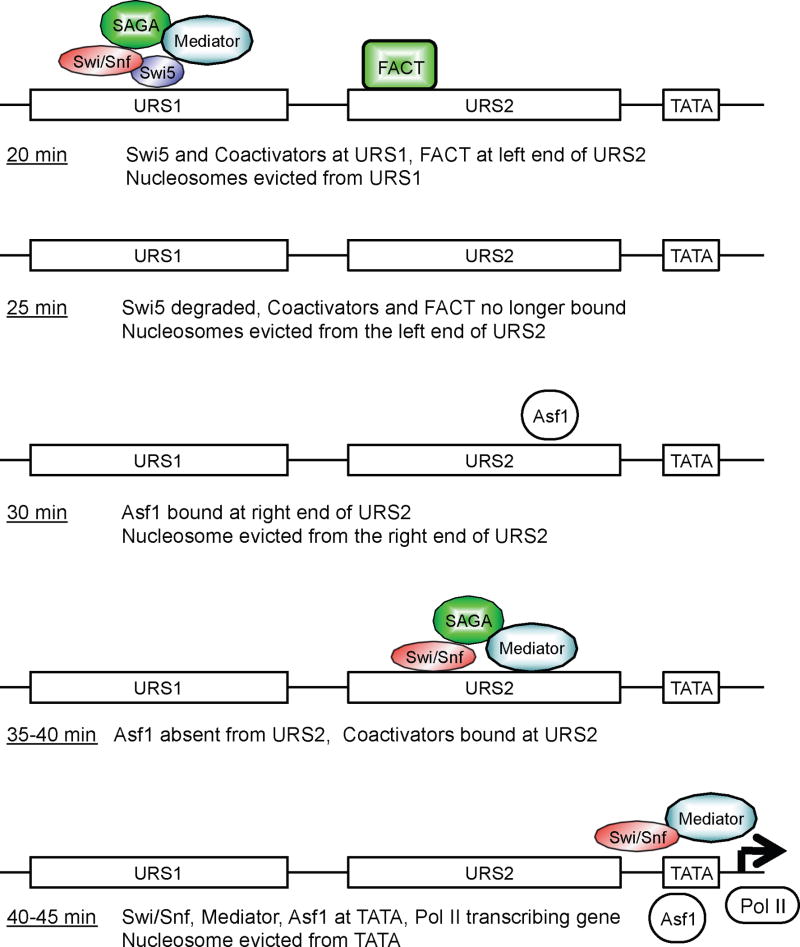
Transcriptional activators and coactivators overcome repression by chromatin, but regulation of chromatin disassembly and coactivator binding to promoters is poorly understood. Activation of the yeast HO gene follows the sequential binding of both sequence-specific DNA-binding proteins and coactivators during the cell cycle. Here, we show that the nucleosome disassembly occurs in waves both along the length of the promoter and during the cell cycle. Different chromatin modifiers are required for chromatin disassembly at different regions of the promoter, with Swi/Snf, the FACT chromatin reorganizer, and the Asf1 histone chaperone each required for nucleosome eviction at distinct promoter regions. FACT and Asf1 both bind to upstream elements of the HO promoter well before the gene is transcribed. The Swi/Snf, SAGA, and Mediator coactivators bind first to the far upstream promoter region and subsequently to a promoter proximal region, and FACT and Asf1 are both required for this coactivator re-recruitment.
Figures
Comment in
-
Keys to open chromatin for transcription activation: FACT and Asf1.Mol Cell. 2009 May 14;34(4):397-9. doi: 10.1016/j.molcel.2009.05.004. Mol Cell. 2009. PMID: 19481517
References
-
- Adkins MW, Howar SR, Tyler JK. Chromatin disassembly mediated by the histone chaperone Asf1 is essential for transcriptional activation of the yeast PHO5 and PHO8 genes. Mol Cell. 2004;14:657–666. - PubMed
-
- Agez M, Chen J, Guerois R, van Heijenoort C, Thuret JY, Mann C, Ochsenbein F. Structure of the histone chaperone ASF1 bound to the histone H3 C-terminal helix and functional insights. Structure. 2007;15:191–199. - PubMed
-
- Barlev NA, Candau R, Wang L, Darpino P, Silverman N, Berger SL. Characterization of physical interactions of the putative transcriptional adaptor, ADA2, with acidic activation domains and TATA-binding protein. J Biol Chem. 1995;270:19337–19344. - PubMed
-
- Berger SL, Pina B, Silverman N, Marcus GA, Agapite J, Regier JL, Triezenberg SJ, Guarente L. Genetic isolation of ADA2: a potential transcriptional adaptor required for function of certain acidic activation domains. Cell. 1992;70:251–265. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
