

L1 retrotransposition occurs mainly in embryogenesis and creates somatic mosaicism
- PMID: 19487571
- PMCID: PMC2701581
- DOI: 10.1101/gad.1803909
L1 retrotransposition occurs mainly in embryogenesis and creates somatic mosaicism
Abstract
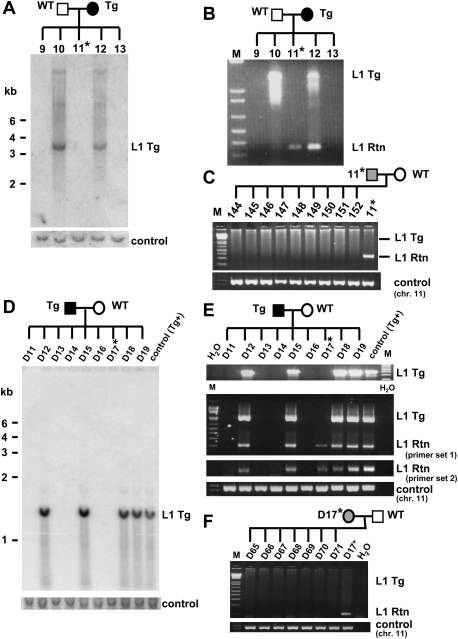
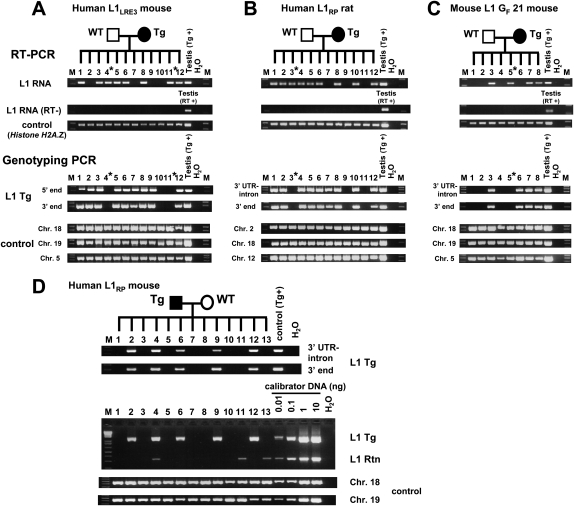
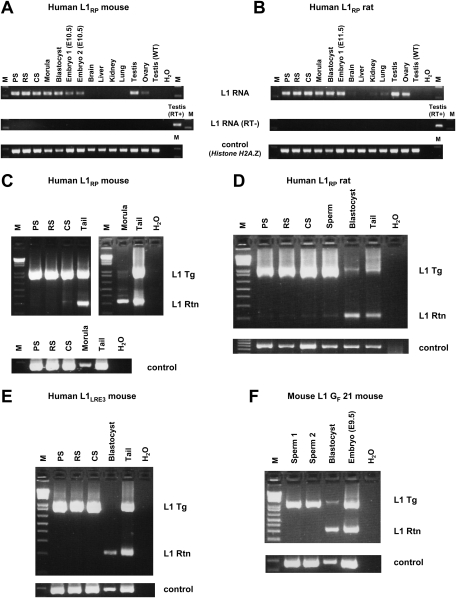
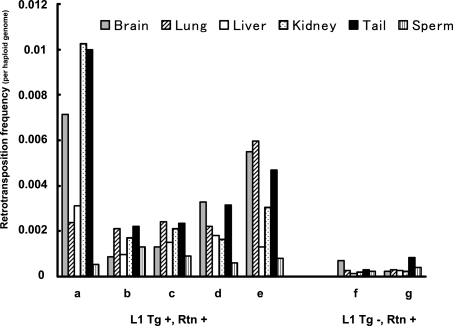
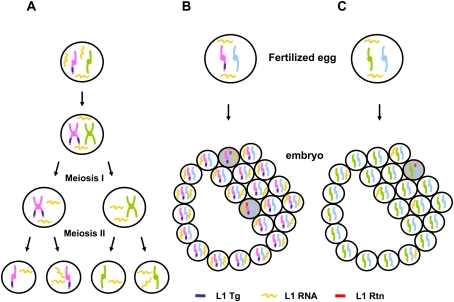
Long Interspersed Element 1 (L1) is a retrotransposon that comprises approximately 17% of the human genome. Despite its abundance in mammalian genomes, relatively little is understood about L1 retrotransposition in vivo. To study the timing and tissue specificity of retrotransposition, we created transgenic mouse and rat models containing human or mouse L1 elements controlled by their endogenous promoters. Here, we demonstrate abundant L1 RNA in both germ cells and embryos. However, the integration events usually occur in embryogenesis rather than in germ cells and are not heritable. We further demonstrate L1 RNA in preimplantation embryos lacking the L1 transgene and L1 somatic retrotransposition events in blastocysts and adults lacking the transgene. Together, these data indicate that L1 RNA transcribed in male or female germ cells can be carried over through fertilization and integrate during embryogenesis, an interesting example of heritability of RNA independent of its encoding DNA. Thus, L1 creates somatic mosaicism during mammalian development, suggesting a role for L1 in carcinogenesis and other disease.
Figures
References
-
- Bourc'his D, Bestor TH. Meiotic catastrophe and retrotransposon reactivation in male germ cells lacking Dnmt3L. Nature. 2004;431:96–99. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases