

The structure of Aquifex aeolicus sulfide:quinone oxidoreductase, a basis to understand sulfide detoxification and respiration
- PMID: 19487671
- PMCID: PMC2689314
- DOI: 10.1073/pnas.0904165106
The structure of Aquifex aeolicus sulfide:quinone oxidoreductase, a basis to understand sulfide detoxification and respiration
Abstract
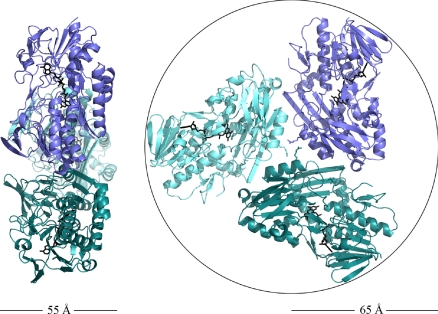
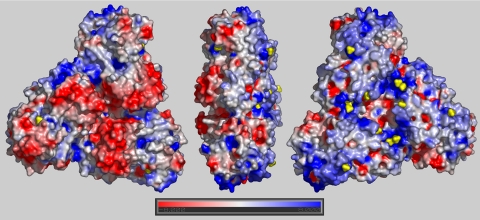
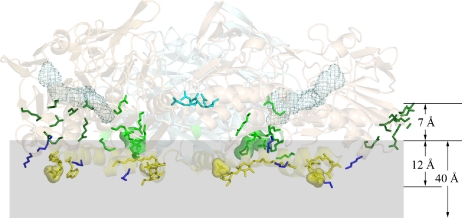
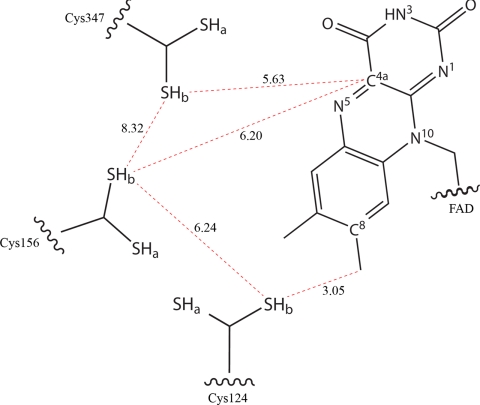
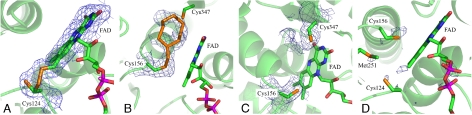
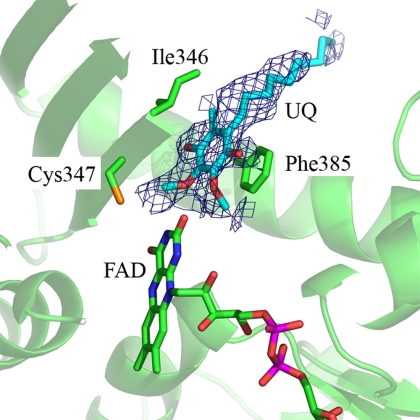
Sulfide:quinone oxidoreductase (SQR) is a flavoprotein with homologues in all domains of life except plants. It plays a physiological role both in sulfide detoxification and in energy transduction. We isolated the protein from native membranes of the hyperthermophilic bacterium Aquifex aeolicus, and we determined its X-ray structure in the "as-purified," substrate-bound, and inhibitor-bound forms at resolutions of 2.3, 2.0, and 2.9 A, respectively. The structure is composed of 2 Rossmann domains and 1 attachment domain, with an overall monomeric architecture typical of disulfide oxidoreductase flavoproteins. A. aeolicus SQR is a surprisingly trimeric, periplasmic integral monotopic membrane protein that inserts about 12 A into the lipidic bilayer through an amphipathic helix-turn-helix tripodal motif. The quinone is located in a channel that extends from the si side of the FAD to the membrane. The quinone ring is sandwiched between the conserved amino acids Phe-385 and Ile-346, and it is possibly protonated upon reduction via Glu-318 and/or neighboring water molecules. Sulfide polymerization occurs on the re side of FAD, where the invariant Cys-156 and Cys-347 appear to be covalently bound to polysulfur fragments. The structure suggests that FAD is covalently linked to the polypeptide in an unusual way, via a disulfide bridge between the 8-methyl group and Cys-124. The applicability of this disulfide bridge for transferring electrons from sulfide to FAD, 2 mechanisms for sulfide polymerization and channeling of the substrate, S(2-), and of the product, S(n), in and out of the active site are discussed.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Kimura H. Hydrogen sulfide as a neuromodulator. Mol Neurobiol. 2002;26:13–19. - PubMed
-
- Shahak Y, Hauska G. Sulfide oxidation from cyanobacteria to humans: Sulfide-quinone oxidoreductase (SQR) In: Hell R, Dahl C, Knaff DB, Leustek T, editors. Advances in Photosynthesis and Respiration. Vol 27. Heidelberg: Springer; 2008. pp. 319–335.
-
- Nübel T, Klughammer C, Huber R, Hauska G, Schütz M. Sulfide:quinone oxidoreductase in membranes of the hyperthermophilic bacterium Aquifex aeolicus (VF5) Arch Microbiol. 2000;173:233–244. - PubMed
-
- Wang R. Two's company, three's a crowd: Can H2S be the third endogenous gaseous transmitter? FASEB J. 2002;16:1792–1798. - PubMed
-
- Griesbeck C, et al. Mechanism of sulfide-quinone reductase investigated using site-directed mutagenesis and sulfur analysis. Biochemistry. 2002;41:11552–11565. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
