

Aberrant epithelial morphology and persistent epidermal growth factor receptor signaling in a mouse model of renal carcinoma
- PMID: 19487675
- PMCID: PMC2700996
- DOI: 10.1073/pnas.0902031106
Aberrant epithelial morphology and persistent epidermal growth factor receptor signaling in a mouse model of renal carcinoma
Abstract
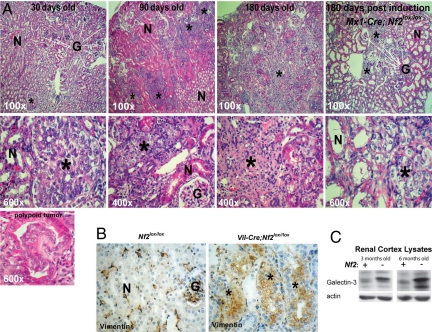
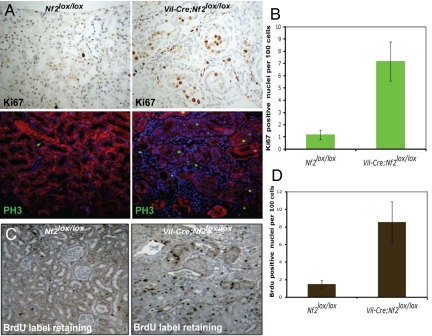
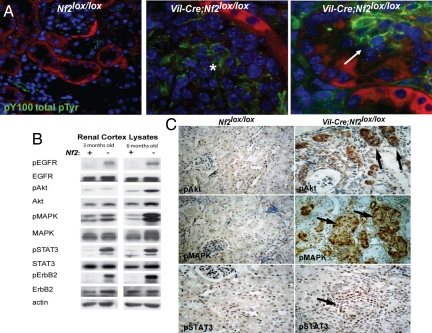
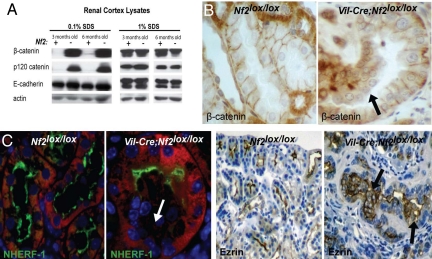
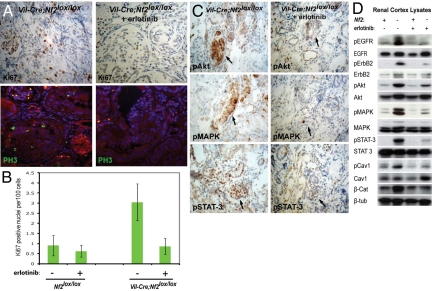
The epidermal growth factor receptor (EGFR) has frequently been implicated in hyperproliferative diseases of renal tubule epithelia. We have shown that the NF2 tumor suppressor Merlin inhibits EGFR internalization and signaling in a cell contact-dependent manner. Interestingly, despite the paucity of recurring mutations in human renal cell carcinoma (RCC), homozygous mutation of the NF2 gene is found in approximately 2% of RCC patient samples in the Sanger COSMIC database. To examine the roles of Merlin and EGFR in kidney tumorigenesis, we generated mice with a targeted deletion of Nf2 in the proximal convoluted epithelium using a Villin-Cre transgene. All of these mice developed intratubular neoplasia by 3 months, which progressed to invasive carcinoma by 6-10 months. Kidneys from these mice demonstrated marked hyperproliferation and a concomitant increase in label-retaining putative progenitor cells. Early lumen-filling lesions in this model exhibited hyperactivation of EGFR signaling, altered solubility of adherens junctions components, and loss of epithelial polarity. Renal cortical epithelial cells derived from either early or late lesions were dependent on EGF for in vitro proliferation and were arrested by pharmacologic inhibition of EGFR or re-expression of Nf2. These cells formed malignant tumors upon s.c. injection into immunocompromised mice before in vitro passage. Treatment of Vil-Cre;Nf2(lox/lox) mice with the EGFR inhibitor erlotinib halted the proliferation of tumor cells. These studies give added credence to the role of EGFR signaling and perhaps Nf2 deficiency in RCC and describe a rare and valuable mouse model for exploring the molecular basis of this disease.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Cohen HT, McGovern FJ. Renal-cell carcinoma. N Engl J Med. 2005;353:2477–2490. - PubMed
-
- Valladares Ayerbes M, et al. Origin of renal cell carcinomas. Clin Transl Oncol. 2008;10:697–712. - PubMed
-
- Iliopoulos O. Molecular biology of renal cell cancer and the identification of therapeutic targets. J Clin Oncol. 2006;24:5593–5600. - PubMed
-
- Motzer RJ, Bander NH, Nanus DM. Renal-cell carcinoma. N Engl J Med. 1996;335:865–875. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
