

doi: 10.1128/MMBR.00041-08.
Signals, regulatory networks, and materials that build and break bacterial biofilms
Affiliations
- PMID: 19487730
- PMCID: PMC2698413
- DOI: 10.1128/MMBR.00041-08
Item in Clipboard
Signals, regulatory networks, and materials that build and break bacterial biofilms
Microbiol Mol Biol Rev.
2009 Jun.
Abstract
Biofilms are communities of microorganisms that live attached to surfaces. Biofilm formation has received much attention in the last decade, as it has become clear that virtually all types of bacteria can form biofilms and that this may be the preferred mode of bacterial existence in nature. Our current understanding of biofilm formation is based on numerous studies of myriad bacterial species. Here, we review a portion of this large body of work including the environmental signals and signaling pathways that regulate biofilm formation, the components of the biofilm matrix, and the mechanisms and regulation of biofilm dispersal.
Figures
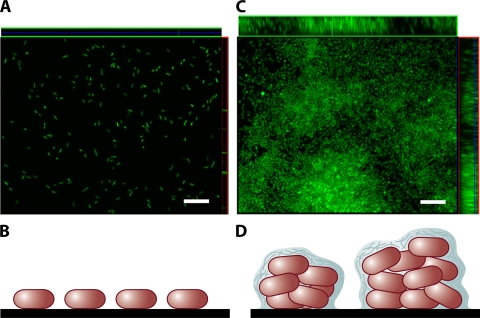
Monolayer and multilayer biofilms. (A and C) Transverse and vertical cross-sections through monolayer (A) and multilayer (C) biofilms of V. cholerae O139. (B and D) Schematic representations of side views of the monolayer (B) and multilayer (D) biofilms. In the monolayer, bacteria are distributed on the surface as a single layer. In the multilayer biofilm, pillars composed of multiple layers of bacteria encased in an extracellular matrix form. Biofilms were grown for 24 h in minimal medium without glucose and in LB broth for monolayer and multilayer biofilms, respectively, as described in reference . Biofilms were stained with Syto 9 and visualized using confocal scanning laser microscopy with an LSM 510 META confocal scanning system. Bars, ∼10 μm.
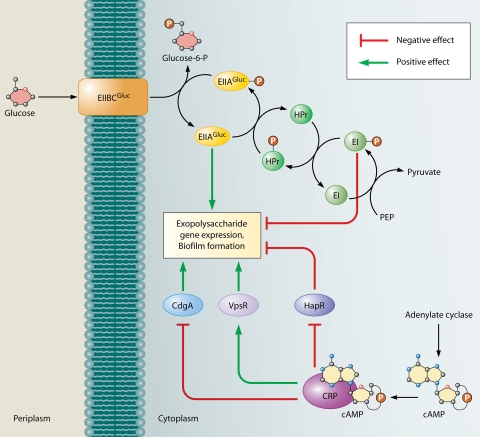
Effect of glucose transport and catabolite repression on V. cholerae biofilm formation. The PTS for glucose regulates biofilm formation in V. cholerae. Unphosphorylated EIIAGluc, which signals the presence of glucose in the environment, leads to activation of biofilm formation, whereas, phosphorylated EI, which signals the absence of glucose in the environment, leads to its repression. The cAMP-CRP complex has been shown to both activate and repress biofilm formation. The activation occurs as a result of negative regulation of the biofilm repressor HapR and positive regulation of the biofilm activator VpsR. The repression is due to negative regulation of the DGC CdgA, an activator of biofilm formation.
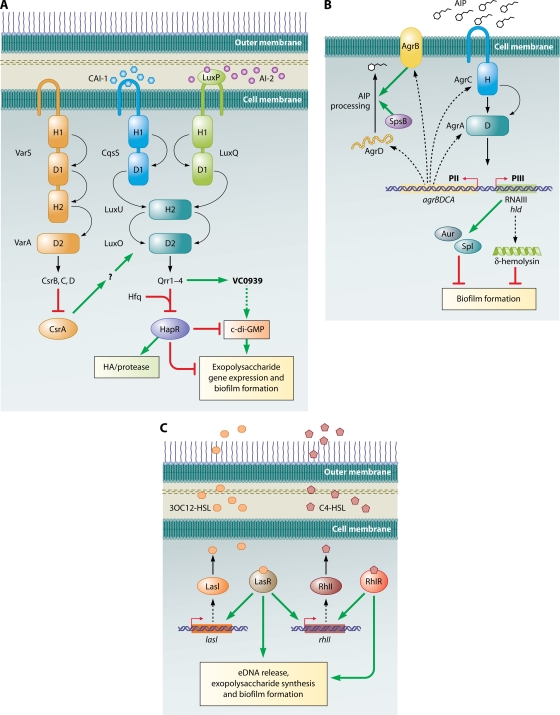
Quorum-sensing circuits and biofilm formation. (A) V. cholerae. Three quorum-sensing circuits converge on HapR to regulate biofilm formation. A HapR-independent quorum-sensing pathway involving Qrr1 to -4 and VC0939, which encodes a GGDEF family protein, has also been identified. This protein is likely to be a DGC that makes c-di-GMP, which is a positive activator of biofilm formation. HapR inhibits biofilm formation via multiple pathways, one of which is by indirectly decreasing c-di-GMP concentrations in the cell. Curved arrows denote the flow of phosphate under low-cell-density conditions. Phosphate flow is reversed at high density. H and D refer to the histidine and aspartate residues, respectively, which accept and shuttle the phosphoryl group. Dotted lines denote hypothesized effects. The question mark refers to a hypothesized intermediate effector in the pathway that has not been identified. (Adapted from reference with permission of Blackwell Publishing Ltd.) (B) S. aureus: the Agr quorum-sensing pathway. A TCS composed of the histidine kinase AgrC and the response regulator AgrA responds to the presence of AIP. Phosphorylated AgrA activates transcription of the divergent PII and PIII operons. The PII operon encodes the machinery to synthesize, process, and detect AIP, while the PIII operon encodes RNAIII, the major effector of the quorum-sensing response. RNAIII regulates numerous downstream genes, two of which encode Aur and Spl proteases that are negative effectors of biofilm formation. The RNAIII transcript also encodes δ-hemolysin, which also inhibits biofilm formation. Curved arrows denote the flow of phosphate under high-cell-density conditions. H and D refer to the histidine and aspartate residues, respectively, which accept and shuttle the phosphoryl group. Broken lines connect the genes to their gene products. (C) P. aeruginosa. Las and Rhl pathways regulate quorum-sensing responses. The Rhl system is under the control of the Las system. LasR and RhlR, in the presence of their cognate autoinducers, activate a large number of genes, among which are those involved in exopolysaccharide production, eDNA, and biofilm formation. Broken lines connect the genes to their gene products.
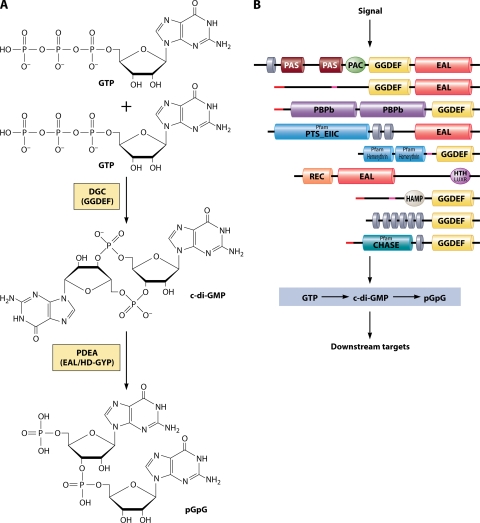
c-di-GMP. (A) Synthesis and breakdown of c-di-GMP. c-di-GMP is synthesized by DGCs containing GGDEF domains from two GTP molecules and broken down to pGpG by PDEAs (containing either EAL or HD-GYP domains). (B) GGDEF and EAL/HD-GYP superfamily proteins are modular and diverse. Proteins that belong to this superfamily contain a variety of sensory domains that are likely to regulate the activity of the enzymatic domains based on the input signals. Shown here are a number of GGDEF/EAL proteins from V. cholerae that have different domain architectures (top to bottom, VC0072, VC0658, VC1067, VC1211, VC1216, VC1652, VC1370, VC1372, and VC1376). Sequences were obtained from http://cmr.jcvi.org . Domain architecture was analyzed using SMART (Simple Modular Architecture ResearchTool) (http://smart.embl-heidelberg.de ) (283). Abbreviations: CHASE, cyclase/histidine kinase-associated sensory domain; GAF, domain present in phytochromes and cGMP-specific phosphodiesterases; HAMP, histidine kinases, adenylyl cyclases, methyl binding proteins, phosphatase domain; hemerythrin, hemerythrin HHE cation binding domain; HTH LUXR, helix-turn-helix, Lux regulon, PAS/PAC, Per (periodic clock protein), Arnt (aryl hydrocarbon receptor nuclear translocator protein), and Sim (single-minded protein) domain; PBPb, bacterial periplasmic substrate binding proteins; PTS_EIIC, PTS, EIIC; Rec, CheY-homologous receiver domain. Gray disks denote predicted transmembrane domains. Red lines denote predicted signal sequences. Pink lines denote segments of low compositional complexity. Pfam, Protein family database (http://pfam.sanger.ac.uk ).
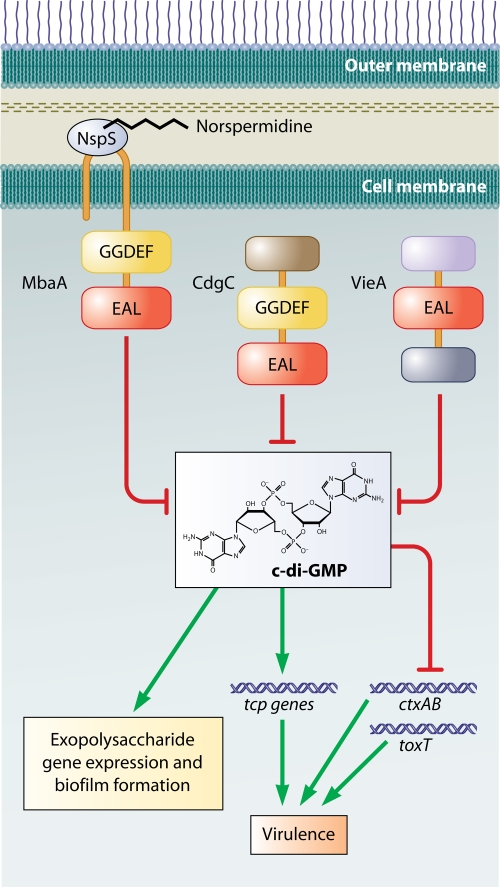
c-di-GMP regulation of virulence and biofilm formation in V. cholerae. MbaA is a predicted c-di-GMP phosphodiesterase. Association of MbaA with NspS is thought to inhibit its activity. Hypothesized binding of norspermidine to NspS is thought to increase this inhibition. CdgC is a phosphodiesterase that decreases c-di-GMP pools in the cell. Deletion of cdgC leads to an increase in intracellular c-di-GMP and an accompanying increase in the transcription of some of the genes in the tcp operon, which is required for virulence. VieA is also a phosphodiesterase which positively effects transcription of the virulence genes ctxAB and toxT via its negative effect on c-di-GMP.
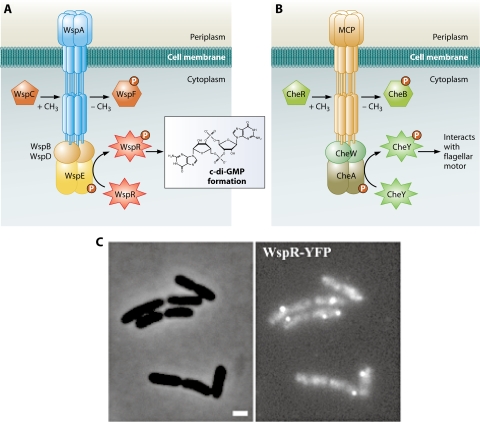
Regulation of intracellular c-di-GMP by a chemotaxis-like signaling system in P. aeruginosa. (A) Predicted organization of the Wsp signaling system. Wsp proteins regulate the c-di-GMP concentrations based on an as-yet-unidentified signal. The signal is predicted to be detected by the methyl-accepting chemotaxis protein homolog WspA; WspE autophosphorylates and transfers the signal to WspR. Phosphorylated WspR has increased DGC activity. WspA is coupled to WspE via WspB and -D. The methylation state of WspA is determined by opposing activities of the methyltransferase homolog WspC and the methylesterase homolog WspF. Demethylation of the receptor by WspF is thought to be involved in adaptation to the signal. (B) Predicted organization of the Che signaling system. The chemotaxis signaling pathway responds to external attractant and repellent molecules and results in swimming toward or away from these molecules, respectively. The output of the system is CheY, which in its phosphorylated form interacts with the flagellar switch proteins to determine the direction of flagellar rotation. (C) Clustering of WspR in the cell. The phase-contrast (left) and fluorescence (right) images of cells expressing a WspR translational fusion to yellow fluorescent protein (WspR-YFP) are shown. Notice clusters of fluorescence, indicating clustering of WspR-YFP. Bar, 1 μm. (Adapted from reference with permission of Blackwell Publishing Ltd.)
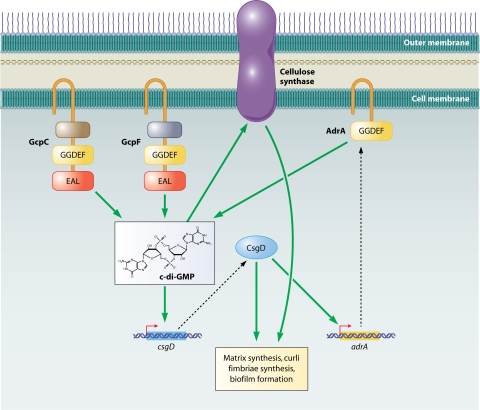
Regulation of biofilm formation in S. Typhimurium by DGCs. Various diguanylated cyclases act both upstream and downstream in the signaling cascade that regulates biofilm formation. GcpC and -F positively affect CsgD, which is the main transcriptional activator of curli biosynthesis genes that are required for biofilm formation as well as adrA. AdrA is also a DGC. c-di-GMP then activates cellulose synthase leading cellulose production, which is part of the matrix of the biofilms made by S. Typhimurium. Broken lines connect the genes to their gene products.
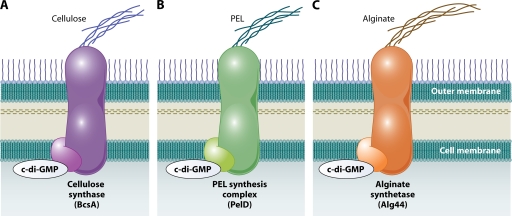
Regulation of exopolysaccharide synthesis machinery by c-di-GMP. Binding of c-di-GMP to both hypothesized and known components of the exopolysaccharide synthesis machinery has been shown for cellulose synthase (A), the PEL synthesis complex (B), and alginate synthetase (C). Activation of these enzymes by c-di-GMP could be a common mechanism of regulating exopolysaccharide synthesis by this second messenger.
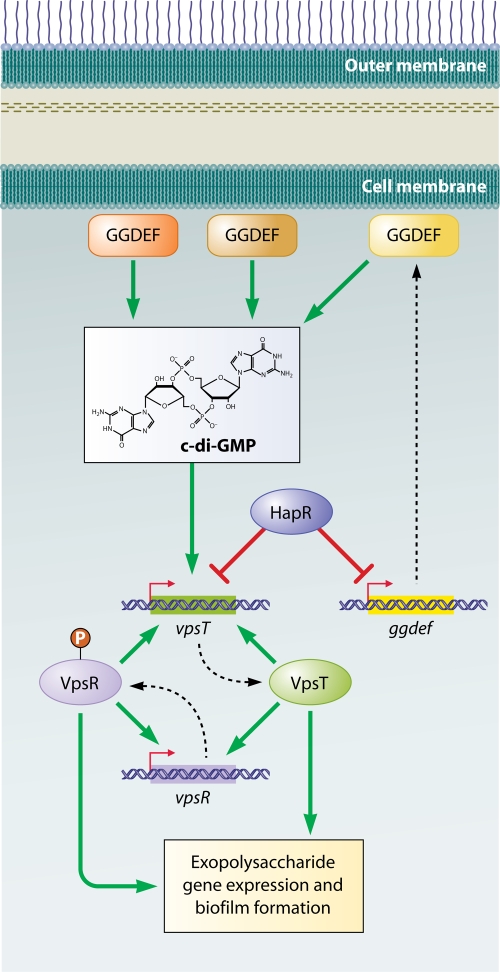
Two-component signaling pathways regulating biofilm formation in V. cholerae. The main regulator of exopolysaccharide gene expression, VpsR, is a two-component response regulator. It also activates its own transcription as well as that of vpsT, which encodes the second two-component response regulator involved in biofilm formation. VpsT also activates its own transcription as well as that of vpsR. vpsT gene transcription is activated by increased c-di-GMP levels and repressed by HapR. HapR also represses transcription of a number of genes encoding GGDEF proteins. Broken lines connect the genes to their gene products.
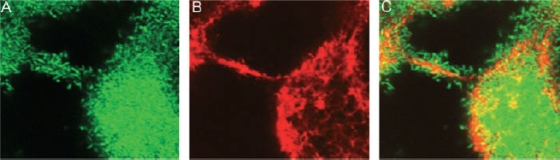
eDNA in the biofilm matrix. Horizontal optical sections in a 2-day-old biofilm formed by green fluorescent protein-tagged P. aeruginosa stained with the DNA binding dye DDAO [7-hydroxy-9H-(1,3-dichloro-9,9-dimethyl acridin-2-one)] are shown. The images show the green fluorescent bacteria (A), the red fluorescent eDNA (B), and an overlay of the two (C). (Reprinted from reference with permission of Blackwell Publishing Ltd.)
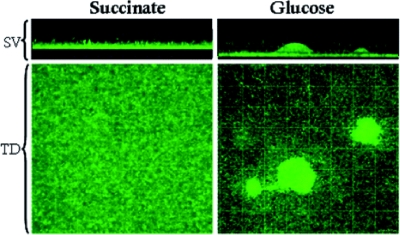
Flat versus irregular topology. Flow cell biofilms of green fluorescent protein-tagged P. aeruginosa grown with different carbon sources are shown. These images were acquired 48 h after inoculation of the system. Biofilms grown with succinate have a flat topology characterized by high surface coverage and low height; those grown with glucose have an irregular topology characterized by low surface coverage and dispersed pillars of bacteria. SV, side view (xz plane); TD, top-down view (xy plane). Gridlines are spaced 20.1 μm apart. (Reprinted from reference with permission of Blackwell Publishing Ltd.)

Quorum-sensing regulation of dispersal. Confocal scanning laser microscopy reconstructions of S. aureus biofilms are shown. The growth medium was supplemented on day 3 with AIP-1 (A) or AIP-1 and the serine protease inhibitor phenylmethylsulfonyl fluoride (PMSF) as indicated (B to D). aur, aureolysin. (Reprinted from reference with permission.)
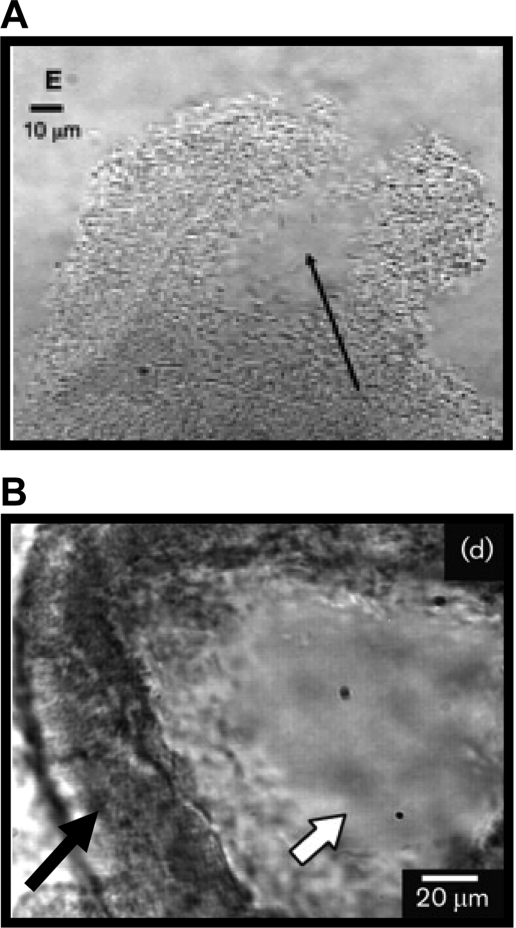
Induction of motility. (A) Cells have evacuated the pillars, leaving a hollow interior (arrow). (Reprinted from reference with permission.) (B) Motile cells inside a wall of stationary cells. The white arrow points to the inside of the pillars where cells have gained motility. Motility is indicated by the blur in the image. The black arrow points to the “walls” of the pillars that are formed by cells that show no motility (see also the first movie in the supplemental material of reference 258). (Reprinted from reference with permission of the publisher.)
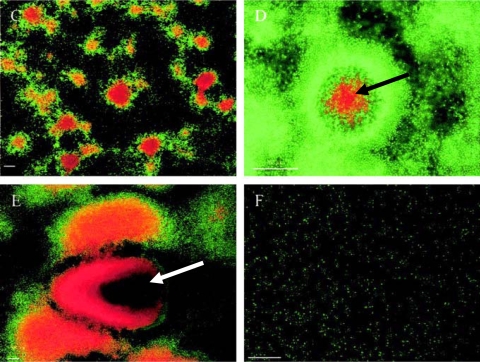
Cell death and cell lysis in biofilm dispersal, showing biofilm development and cell death of the P. tunicata wild-type strain. Biofilms were stained with the BacLight Live/Dead bacterial viability kit. Red propidium iodide-stained cells have a compromised cell membrane and are dead. Time points after inoculation are shown as follows: (C) 48 h; (D) 72 h; (E) 144 h; (F) 168 h. Cell death can be observed at 48 h, and cell lysis (arrow in panel D) and extensive cell death (arrow in panel E) are seen at 144 h, prior to complete dispersal of the biofilm at 168 h. Bars, 50 μm. (Reprinted from reference with permission.)
References
-
- Aldridge, P., and U. Jenal. 1999. Cell cycle-dependent degradation of a flagellar motor component requires a novel-type response regulator. Mol. Microbiol. 32379-391. - PubMed
-
- Allesen-Holm, M., K. B. Barken, L. Yang, M. Klausen, J. S. Webb, S. Kjelleberg, S. Molin, M. Givskov, and T. Tolker-Nielsen. 2006. A characterization of DNA release in Pseudomonas aeruginosa cultures and biofilms. Mol. Microbiol. 591114-1128. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
