

RNA polymerase active center: the molecular engine of transcription
- PMID: 19489723
- PMCID: PMC2929140
- DOI: 10.1146/annurev.biochem.76.052705.164655
RNA polymerase active center: the molecular engine of transcription
Abstract
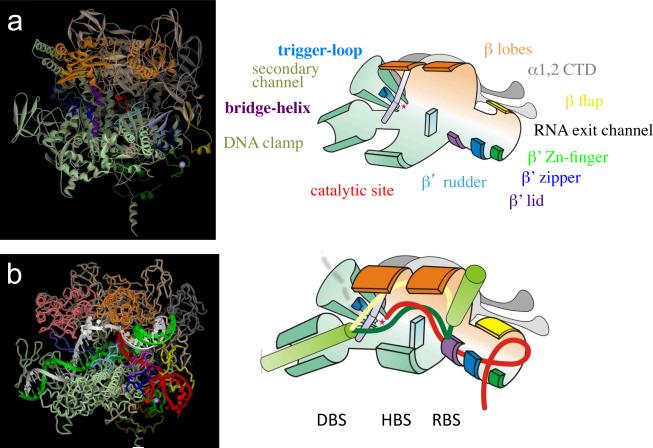
RNA polymerase (RNAP) is a complex molecular machine that governs gene expression and its regulation in all cellular organisms. To accomplish its function of accurately producing a full-length RNA copy of a gene, RNAP performs a plethora of chemical reactions and undergoes multiple conformational changes in response to cellular conditions. At the heart of this machine is the active center, the engine, which is composed of distinct fixed and moving parts that serve as the ultimate acceptor of regulatory signals and as the target of inhibitory drugs. Recent advances in the structural and biochemical characterization of RNAP explain the active center at the atomic level and enable new approaches to understanding the entire transcription mechanism, its exceptional fidelity and control.
Figures




References
-
- Adelman K, et al. Molecular mechanism of transcription inhibition by peptide antibiotic Microcin J25. Mol. Cell. 2004;14:753–762. - PubMed
-
- Artsimovitch I, et al. Allosteric modulation of the RNA polymerase catalytic reaction is an essential component of transcription control by rifamycins. Cell. 2005;122:351–63. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
