

Lipid-dependent membrane protein topogenesis
- PMID: 19489728
- PMCID: PMC3033430
- DOI: 10.1146/annurev.biochem.77.060806.091251
Lipid-dependent membrane protein topogenesis
Abstract
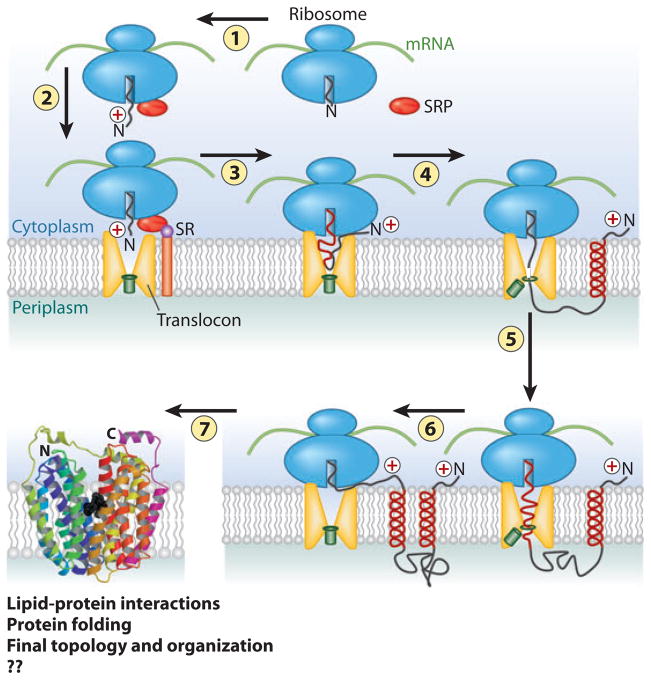
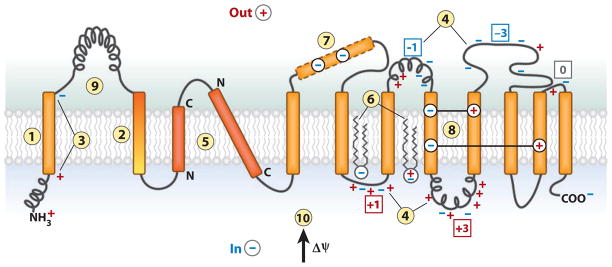
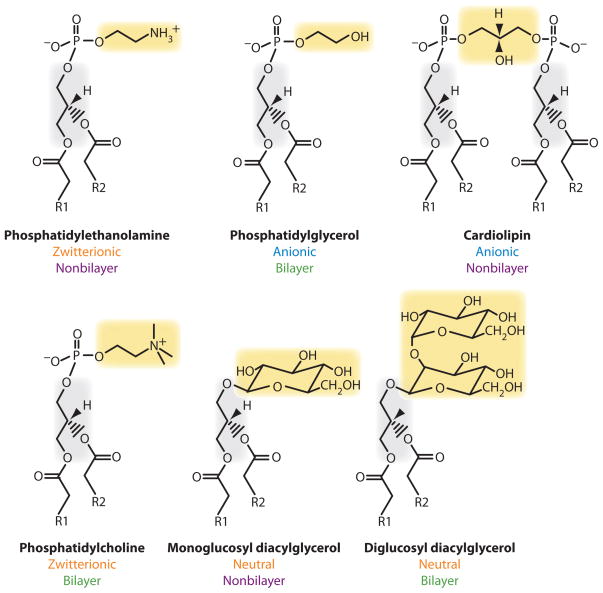
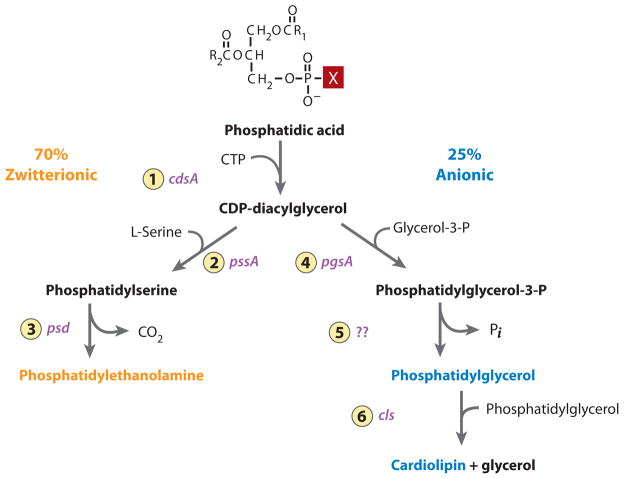
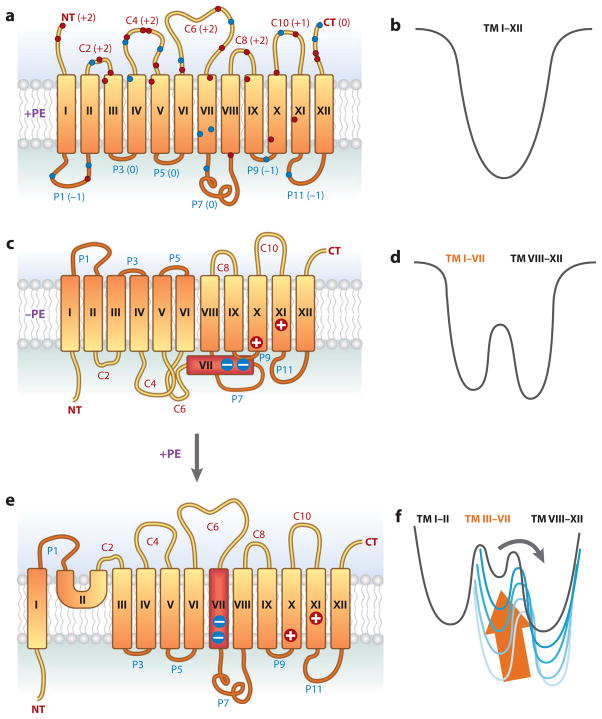
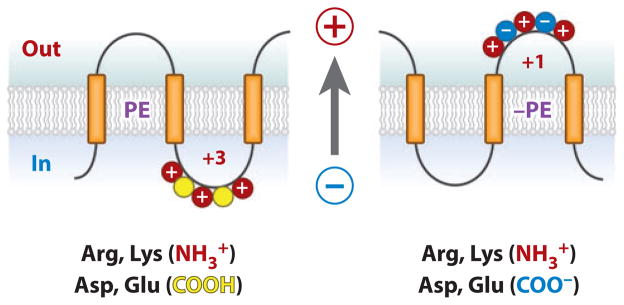
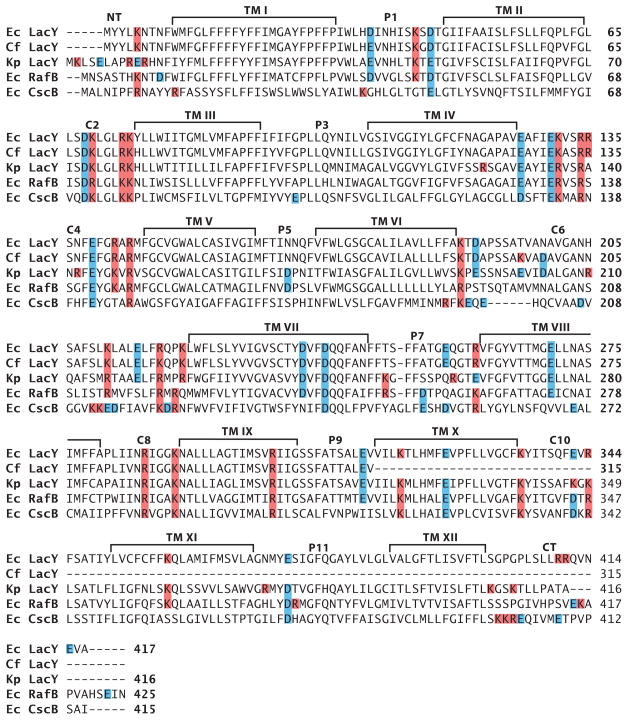
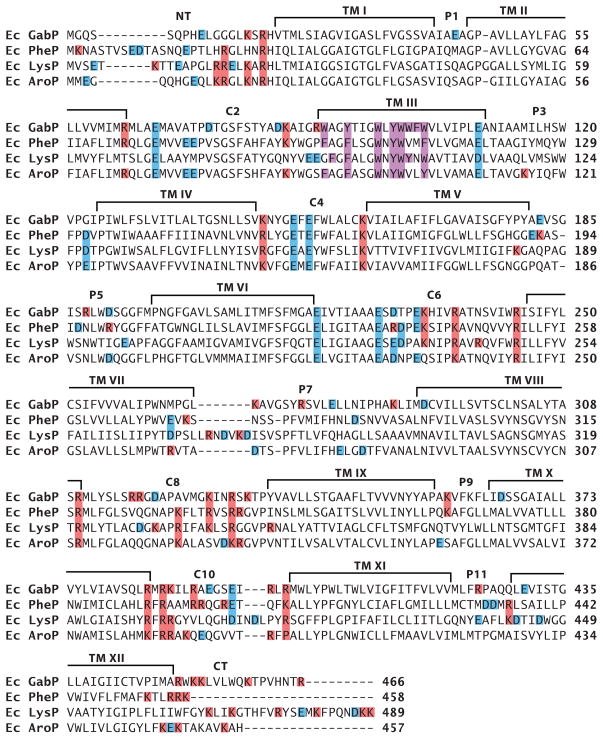
The topology of polytopic membrane proteins is determined by topogenic sequences in the protein, protein-translocon interactions, and interactions during folding within the protein and between the protein and the lipid environment. Orientation of transmembrane domains is dependent on membrane phospholipid composition during initial assembly as well as on changes in lipid composition postassembly. The membrane translocation potential of negative amino acids working in opposition to the positive-inside rule is largely dampened by the normal presence of phosphatidylethanolamine, thus explaining the dominance of positive residues as retention signals. Phosphatidylethanolamine provides the appropriate charge density that permits the membrane surface to maintain a charge balance between membrane translocation and retention signals and also allows the presence of negative residues in the cytoplasmic face of proteins for other purposes.
Figures
References
-
- Driessen AJ, Nouwen N. Protein translocation across the bacterial cytoplasmic membrane. Annu Rev Biochem. 2008;77:643–67. - PubMed
-
- Rapoport TA. Protein translocation across the eukaryotic endoplasmic reticulum and bacterial plasma membranes. Nature. 2007;450:663–69. - PubMed
-
- Pitonzo D, Skach WR. Molecular mechanisms of aquaporin biogenesis by the endoplasmic reticulum Sec61 translocon. Biochim Biophys Acta. 2006;1758:976–88. - PubMed
-
- von Heijne G. Membrane-protein topology. Nat Rev Mol Cell Biol. 2006;7:909–18. - PubMed
-
- Sadlish H, Pitonzo D, Johnson AE, Skach WR. Sequential triage of transmembrane segments by Sec61α during biogenesis of a native multispanning membrane protein. Nat Struct Mol Biol. 2005;12:870–88. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
