

Input-specific plasticity of N-methyl-D-aspartate receptor-mediated synaptic responses in neonatal rat motoneurons
- PMID: 19490018
- PMCID: PMC2931593
- DOI: 10.1111/j.1460-9568.2009.06769.x
Input-specific plasticity of N-methyl-D-aspartate receptor-mediated synaptic responses in neonatal rat motoneurons
Abstract
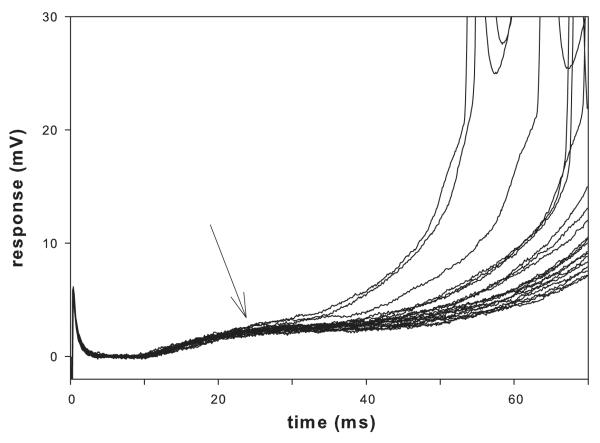
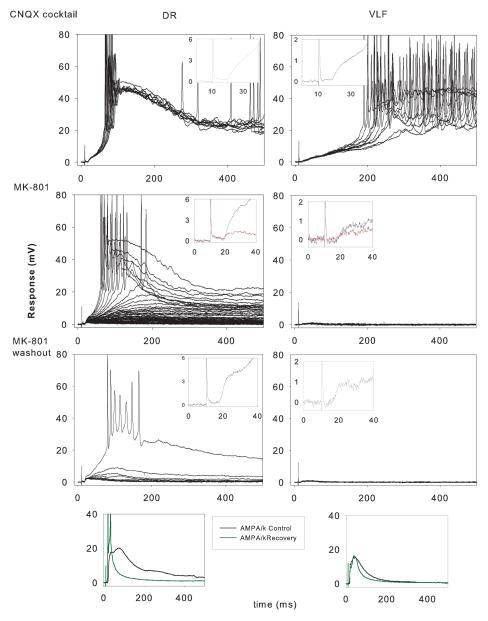
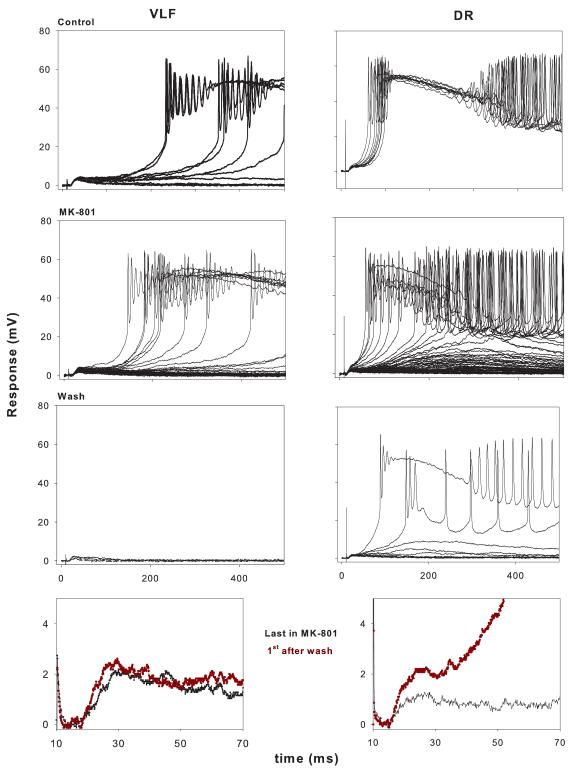
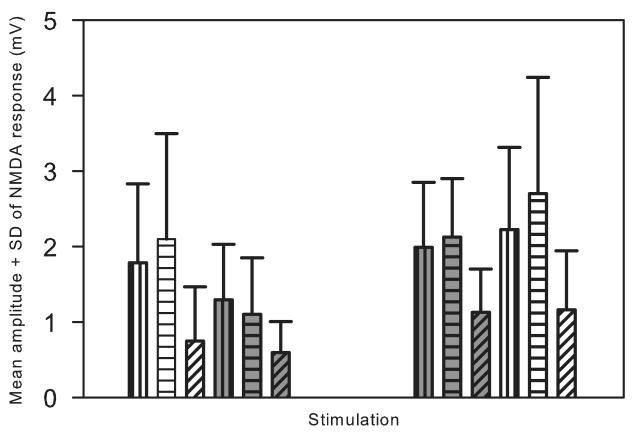
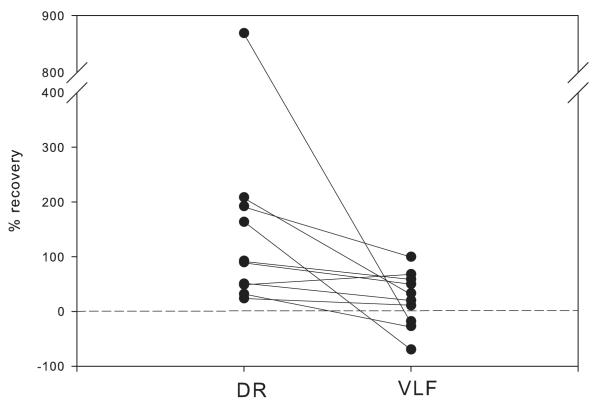
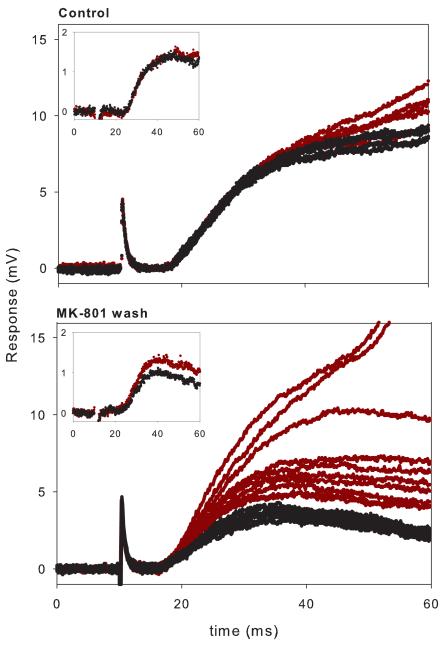
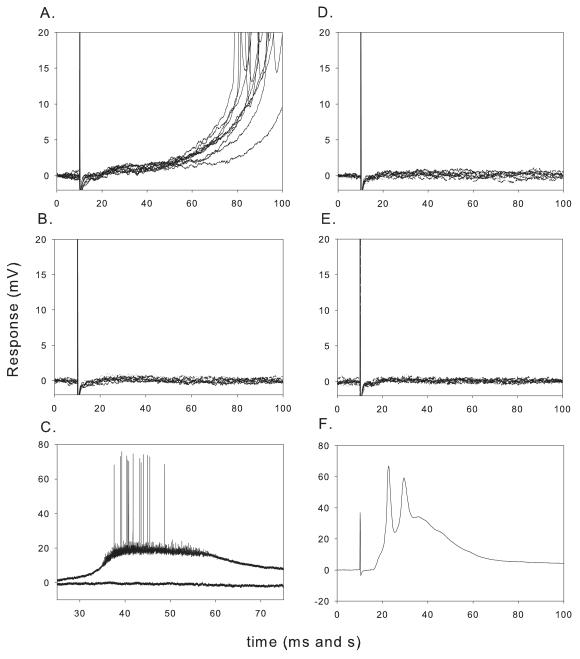
Lumbar motoneurons can be activated monosynaptically by two glutamatergic synaptic inputs: the segmental dorsal root (DR) and the descending ventrolateral funiculus (VLF). To determine whether their N-methyl-d-aspartate (NMDA) receptors are independent, we used (5R,10S)-(+)-5-methyl-10,11-dihydro-5H-dibenzo[a,d]cyclohepten-5,10-imine-hydrogen-maleate (MK-801), known to induce a use-dependent irreversible block of NMDA receptors (NMDARs). In the presence of MK-801 (in bath) and non-NMDA antagonists (in bath, to isolate NMDARs pharmacologically), we first stimulated the DR. After MK-801 blockade of DR synaptic input, the VLF was stimulated. Its response was found to be not significantly different from its control value, suggesting that the DR stimulus activated very few, if any, receptors also activated by VLF stimulation. Similar findings were obtained if the stimulation order was reversed. Both inputs also elicited a polysynaptic NMDAR-mediated response. Evoking the DR polysynaptic response in the presence of MK-801 eliminated the corresponding VLF response; the reverse did not occur. Surprisingly, when MK-801 was washed from the bath, both the DR and the VLF responses could recover, although the recovery of the DR monosynaptic and polysynaptic responses was reliably greater than those associated with the VLF. Recovery was prevented if extrasynaptic receptors were activated by bath-applied NMDA in the presence of MK-801, consistent with the possibility that recovery was due to movement of extrasynaptic receptors into parts of the membrane accessible to transmitter released by DR and VLF stimulation. These novel findings suggest that segmental glutamatergic inputs to motoneurons are more susceptible to plastic changes than those from central nervous system white matter inputs at this developmental stage.
Figures
Similar articles
-
Differential NR2B subunit expression at dorsal root and ventrolateral funiculus synapses on lumbar motoneurons of neonatal rat.Neuroscience. 2010 Mar 17;166(2):730-7. doi: 10.1016/j.neuroscience.2009.12.025. Epub 2009 Dec 16. Neuroscience. 2010. PMID: 20018230 Free PMC article.
-
Removal of NMDA receptor Mg(2+) block extends the action of NT-3 on synaptic transmission in neonatal rat motoneurons.J Neurophysiol. 2001 Jul;86(1):123-9. doi: 10.1152/jn.2001.86.1.123. J Neurophysiol. 2001. PMID: 11431494
-
Acute modulation of synaptic transmission to motoneurons by BDNF in the neonatal rat spinal cord.Eur J Neurosci. 2001 Dec;14(11):1800-8. doi: 10.1046/j.0953-816x.2001.01811.x. Eur J Neurosci. 2001. PMID: 11860475
-
Synaptic plasticity of NMDA receptors: mechanisms and functional implications.Curr Opin Neurobiol. 2012 Jun;22(3):496-508. doi: 10.1016/j.conb.2012.01.007. Epub 2012 Feb 8. Curr Opin Neurobiol. 2012. PMID: 22325859 Free PMC article. Review.
-
Unconventional NMDA Receptor Signaling.J Neurosci. 2017 Nov 8;37(45):10800-10807. doi: 10.1523/JNEUROSCI.1825-17.2017. J Neurosci. 2017. PMID: 29118208 Free PMC article. Review.
Cited by
-
Maladaptive spinal plasticity opposes spinal learning and recovery in spinal cord injury.Front Physiol. 2012 Oct 10;3:399. doi: 10.3389/fphys.2012.00399. eCollection 2012. Front Physiol. 2012. PMID: 23087647 Free PMC article.
-
Differential NR2B subunit expression at dorsal root and ventrolateral funiculus synapses on lumbar motoneurons of neonatal rat.Neuroscience. 2010 Mar 17;166(2):730-7. doi: 10.1016/j.neuroscience.2009.12.025. Epub 2009 Dec 16. Neuroscience. 2010. PMID: 20018230 Free PMC article.
-
Modulation of H-reflex responses and frequency-dependent depression by repetitive spinal electromagnetic stimulation: From rats to humans and back to chronic spinal cord injured rats.Eur J Neurosci. 2020 Dec;52(12):4875-4889. doi: 10.1111/ejn.14885. Epub 2020 Jul 12. Eur J Neurosci. 2020. PMID: 32594554 Free PMC article.
References
-
- Aamodt SM, Constantine-Paton M. The role of neural activity in synaptic development and its implications for adult brain function. Adv. Neurol. 1999;79:133–144. - PubMed
-
- Arvanian VL, Bowers WJ, Petruska JC, Manuzon H, Narrow WC, Motin V, Federoff HJ, Mendell LM. Viral delivery of NR2D subunits reduces Mg2+ block of NMDA receptor and restores NT-3-induced potentiation of AMPA/kainate responses in maturing rat motoneurons. J. Neurophysiol. 2004;92:2394–2404. - PubMed
-
- Arvanian VL, Mendell LM. Acute modulation of synaptic transmission to motoneurons by BDNF in the neonatal rat spinal cord. Eur. J. Neurosci. 2001a;14:1800–1808. - PubMed
-
- Arvanian VL, Mendell LM. Acute modulation of synaptic transmission to motoneurons by BDNF in the neonatal rat spinal cord. Eur. J. Neurosci. 2001b;14:1800–1808. - PubMed
-
- Arvanian VL, Motin V, Mendell LM. Comparison of metabotropic glutamate receptor responses at segmental and descending inputs to motoneurons in neonatal rat spinal cord. J. Pharm. Exp. Ther. 2005;312:669–677. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
