

Review
doi: 10.1016/j.cell.2009.05.002.
The tortoise and the hair: slow-cycling cells in the stem cell race
Affiliations
- PMID: 19490891
- PMCID: PMC2716122
- DOI: 10.1016/j.cell.2009.05.002
Item in Clipboard
Review
The tortoise and the hair: slow-cycling cells in the stem cell race
Cell.
.
Abstract
Given the importance of stem cells to adult tissues, it has long been postulated that stem cells divide infrequently to preserve their long-term proliferation potential and to prevent the acquisition of errors during DNA replication. Yet, some stem cells must be able to continually churn out progeny in tissues that rapidly turn over or are subject to sudden injuries or growth spurts. This Review explores the challenges that mammalian stem cells face in balancing the competing demands of proliferation and differentiation in tissues.
Figures
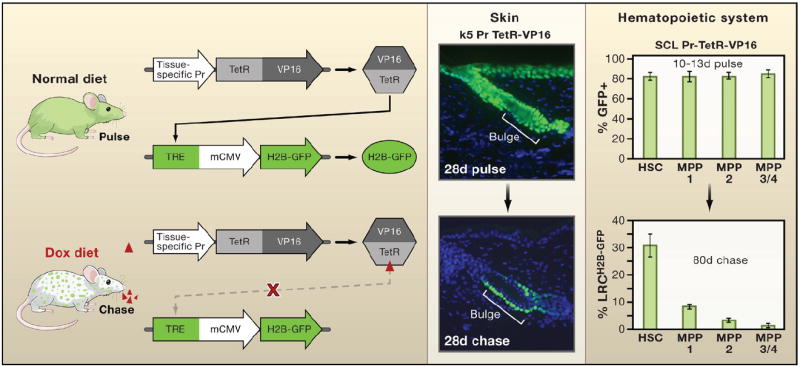
The mating of two parent strains of transgenic mice is needed to make progeny in which expression of a transgene encoding histone H2B-green fluorescent protein (H2B-GFP) can be turned off when tetracycline is added to the animal’s diet. The first parent strain harbors the H2B-GFP transgene under the control of a tetracycline (doxycycline; dox) regulatory element (TRE). The second expresses a transcription factor regulated by tetracycline (TetRVP16) that is under the control of a cell-type-specific promoter. In the first example shown, the keratin 5 (k5) promoter drives expression of TetRVP16 leading to expression of H2B-GFP in the skin epithelium until 4 weeks of age, at which time tetracycline is administered for 4 weeks so that dividing cells dilute out the label and differentiating cells are sloughed from the skin (Tumbar et al., 2004). In the second example, TetRVP16 driven by the promoter of the stem cell leukemia (SCL) gene is used to express H2B-GFP in hematopoietic stem cells (HSCs) and different multipotent progenitors (MPPs) of the hematopoietic system for 10–13 days, after which tetracycline is administered for 80 days to identify the label-retaining cells (LRCs) (Wilson et al., 2008). Schematic adapted from Nowak (2009).
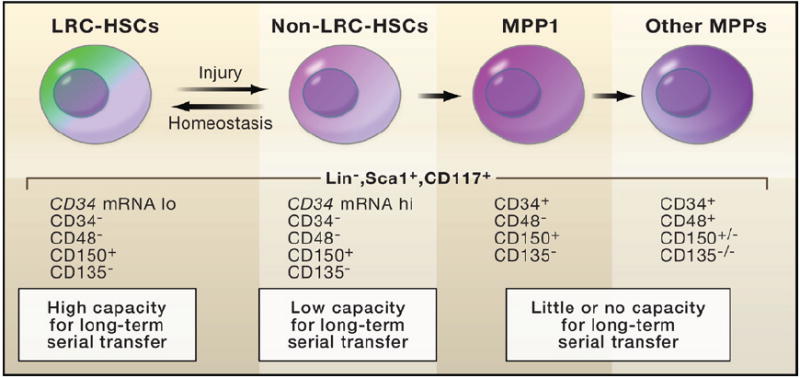
Wilson et al. (2008) coupled the SCL-TetRVP16 pulse-chase experiment (outlined in Figure 1) to fluorescence-activated cell sorting (FACS) in order to purify hematopoietic cell populations. Serial transplant experiments were then used to test the potential of individual hematopoietic cells to reconstitute the bone marrow of X-irradiated mice. The results suggest that cells with the greatest long-term capacity are those that cycle the least frequently. These cells have traditionally been referred to as long-term hematopoietic stem cells (LT-HSCs). They cycle as few as five times per lifetime of the mouse and remain dormant unless challenged by injury or stimuli that induce growth. From these assays, the HSCs that divide slowly but more frequently appear to be the ones that function in normal homeostasis. LRC, label-retaining cells.
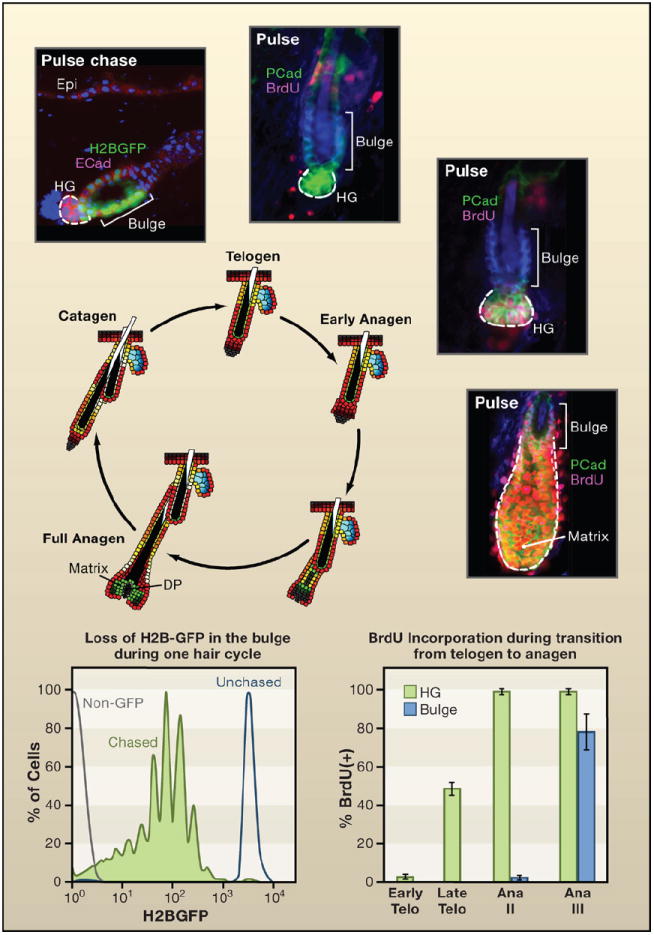
Shown is a schematic of the hair cycle, accompanied by the proliferation state of the stem cell niche. The bulge is a niche for cells that divide relatively infrequently. It is found at the base of the permanent segment of the hair follicle. After the destructive phase (catagen), the hair germ (HG) emerges as a small cluster of cells. The HG is thought to arise from bulge cells that migrate out to “meet” the dermal papilla (DP) that contracts upward (Ito et al., 2004). Histone H2B-GFP pulse-chase experiments show that the upper portion of the HG is label retaining but typically less so than many other bulge cells. HG cells express P-cadherin more strongly than bulge cells (shown). During the resting phase (telogen), both HG and bulge are quiescent. Short BrdU pulses reveal little activity throughout this phase, which can last for many weeks (Greco et al., 2009). Late in telogen and transitioning to early anagen, the HG becomes mobilized prior to the bulge. A few days later (Ana III), the HG has rapidly expanded to form the early matrix of transiently amplifying cells that form the hair shaft and its channel (Greco et al., 2009). At this time, the bulge begins to proliferate. By monitoring reductions in H2B-GFP, the bulge cells divide on average three times per hair cycle, approximately once per week (Waghmare et al., 2008). Thus in contrast to some stem cell niches, such as the intestinal niche, the cycling rates of stem cells are highly sensitive to the relative stage of the hair cycle (Blanpain et al., 2004; Greco et al., 2009; Jaks et al., 2008; Nowak et al., 2008; Waghmare et al., 2008). Schematic and data adapted from Nowak (2009).
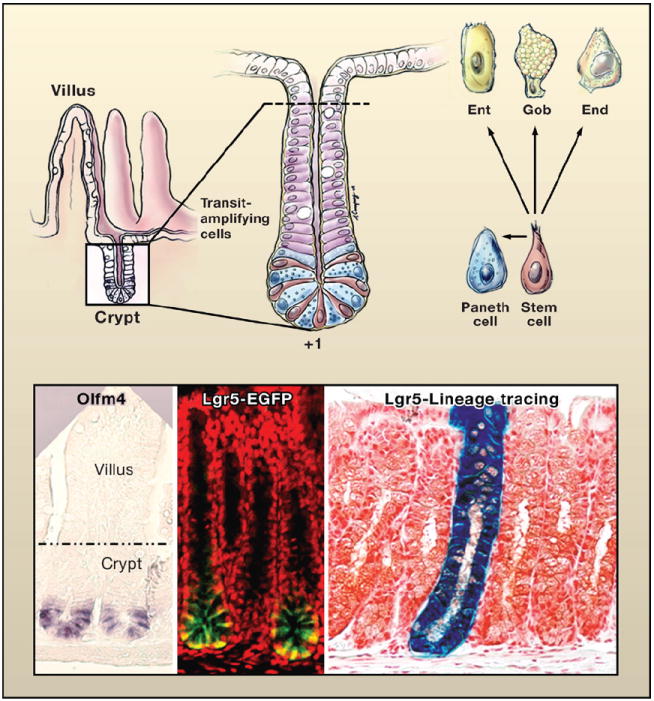
(Top left) Schematic of the crypt-villus architecture of the small intestine. (Top right) Magnified view of the crypt. At the base of the crypt is the putative stem cell zone (Bjerknes and Cheng, 1981; Cheng and Leblond, 1974). This zone contains undifferentiated cells sharing a similar ultrastructure that are interspersed with highly differentiated antibacterial Paneth cells. The undifferentiated columnar cells, purported to be stem cells, extend from a position +1 at the very base of the crypt to +4, marking the boundary between the Paneth cells and the transiently amplifying, rapidly dividing cells that give rise to the terminally differentiating enterocytes (Ent), goblet cells (Gob), and enteroendocrine cells (End) of the villus. Drawing by A. Canapary. (Lower panels) In situ hybridization with a probe for Olfm4, enriched in the stem cell zone (van der Flier et al., 2009). The expression of an Lgr5-EGFP-CreER transgene marks the stem cell zone (Barker et al., 2007). To the right is a lineage tracing of a bitransgenic mouse expressing Lgr5-EGFP-CreER and Rosa26-fl-stop-fl-LacZ. Cells in which the Lgr5 promoter is active are marked by EGFP expression and express tamoxifen-regulatable Cre recombinase. Following activation of Cre, floxed sequences recombine, excising the stop codon and activating LacZ. With time, the entire crypt-villus structure displays β-galactosidase expressing progeny, revealing that individual Lgr5-positive stem cells can regenerate the entire niche (Barker et al., 2007). Images reprinted by permission from Macmillan Publishers Ltd: Nature (Barker et al., 2007), copyright 2007.
References
-
- Barker N, van Es JH, Kuipers J, Kujala P, van den Born M, Cozijnsen M, Haegebarth A, Korving J, Begthel H, Peters PJ, Clevers H. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature. 2007;449:1003–1007. - PubMed
-
- Barker N, Ridgway RA, van Es JH, van de Wetering M, Begthel H, van den Born M, Danenberg E, Clarke AR, Sansom OJ, Clevers H. Crypt stem cells as the cells-of-origin of intestinal cancer. Nature. 2009;457:608–611. - PubMed
-
- Bjerknes M, Cheng H. The stem-cell zone of the small intestinal epithelium. IV. Effects of resecting 30% of the small intestine. Am J Anat. 1981;160:93–103. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
