

The mRNA-destabilizing protein tristetraprolin is suppressed in many cancers, altering tumorigenic phenotypes and patient prognosis
- PMID: 19491267
- PMCID: PMC2724875
- DOI: 10.1158/0008-5472.CAN-08-4238
The mRNA-destabilizing protein tristetraprolin is suppressed in many cancers, altering tumorigenic phenotypes and patient prognosis
Abstract
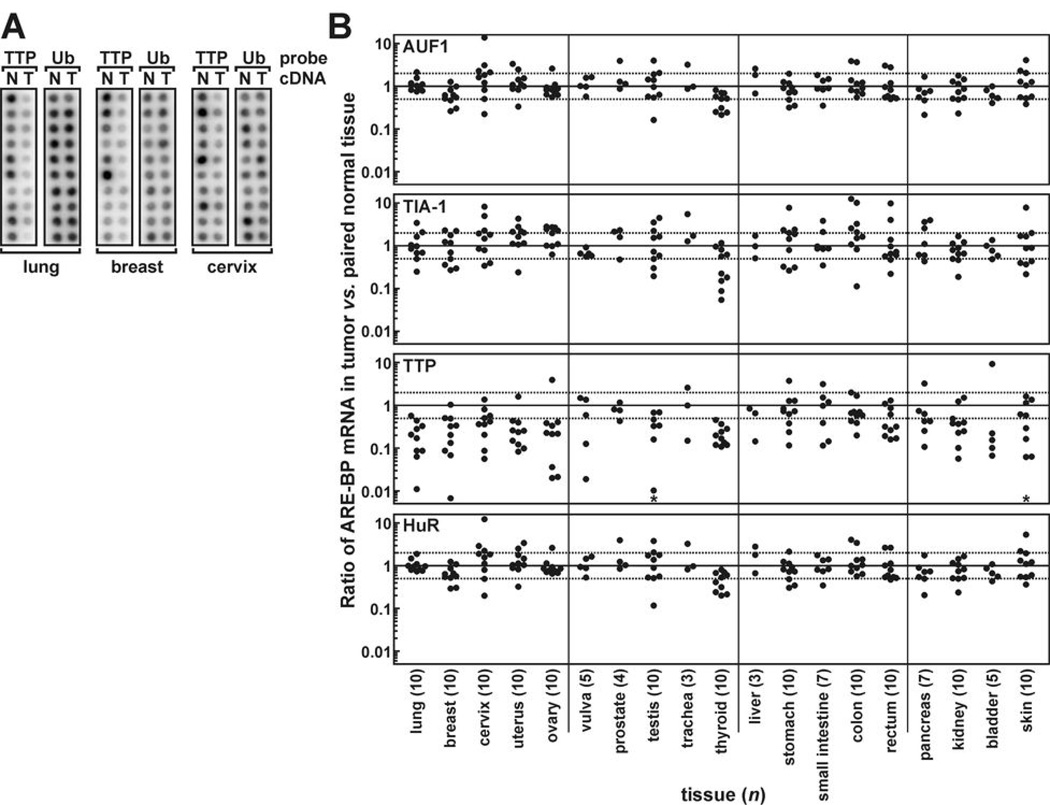
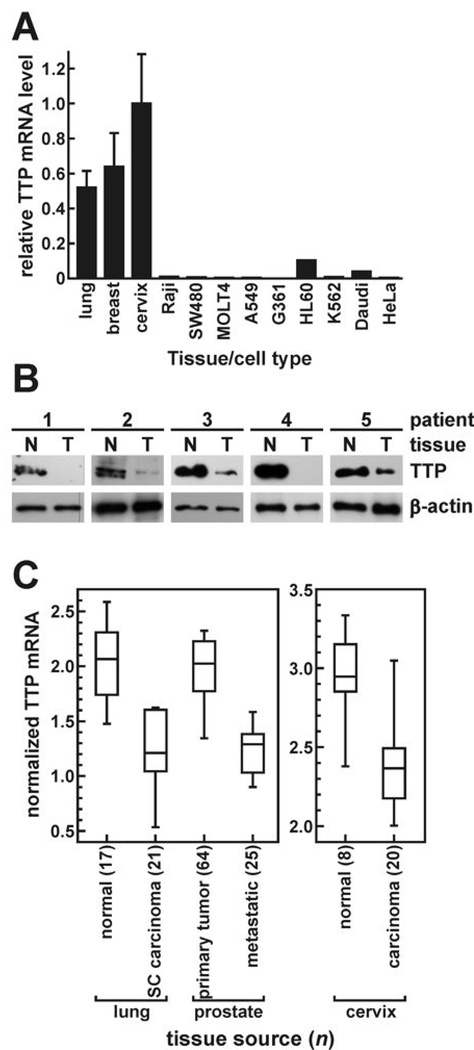
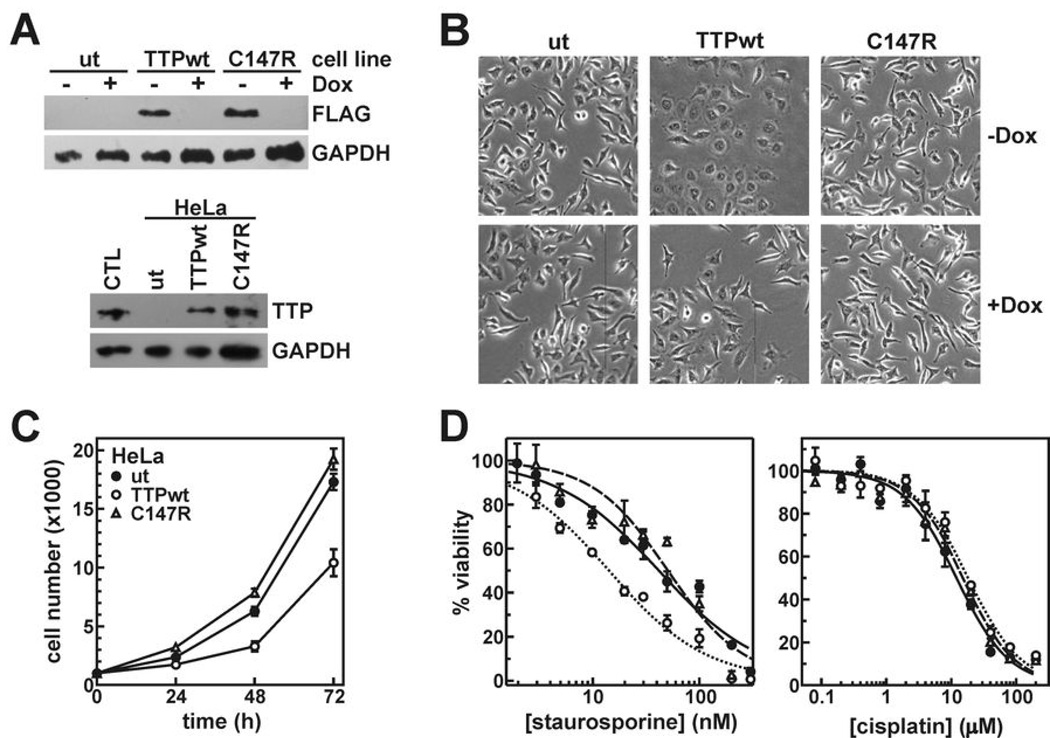
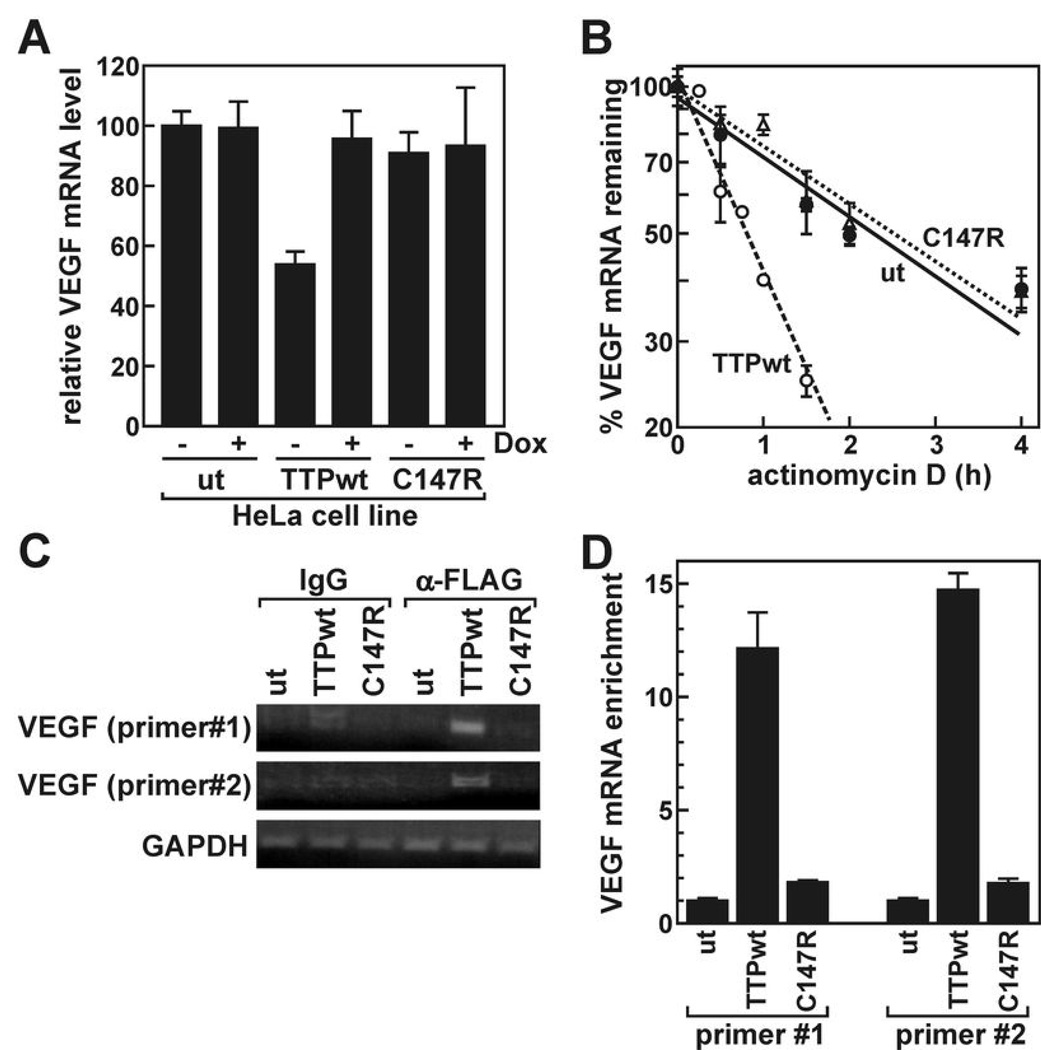
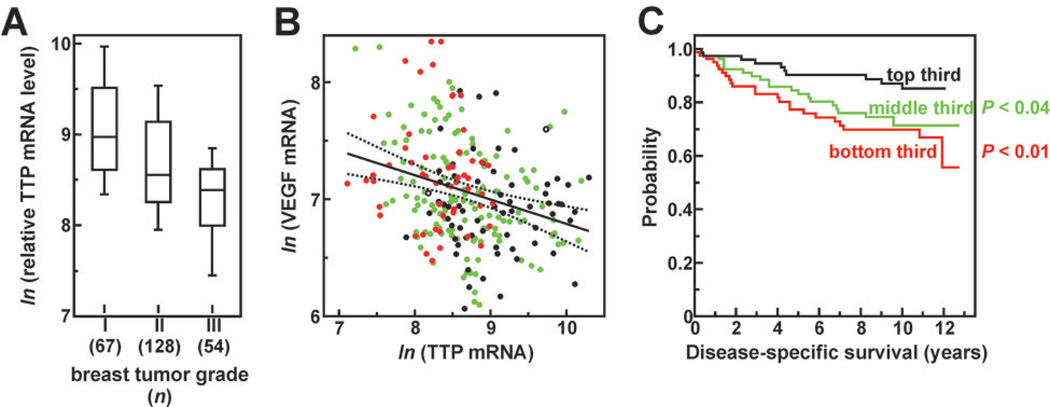
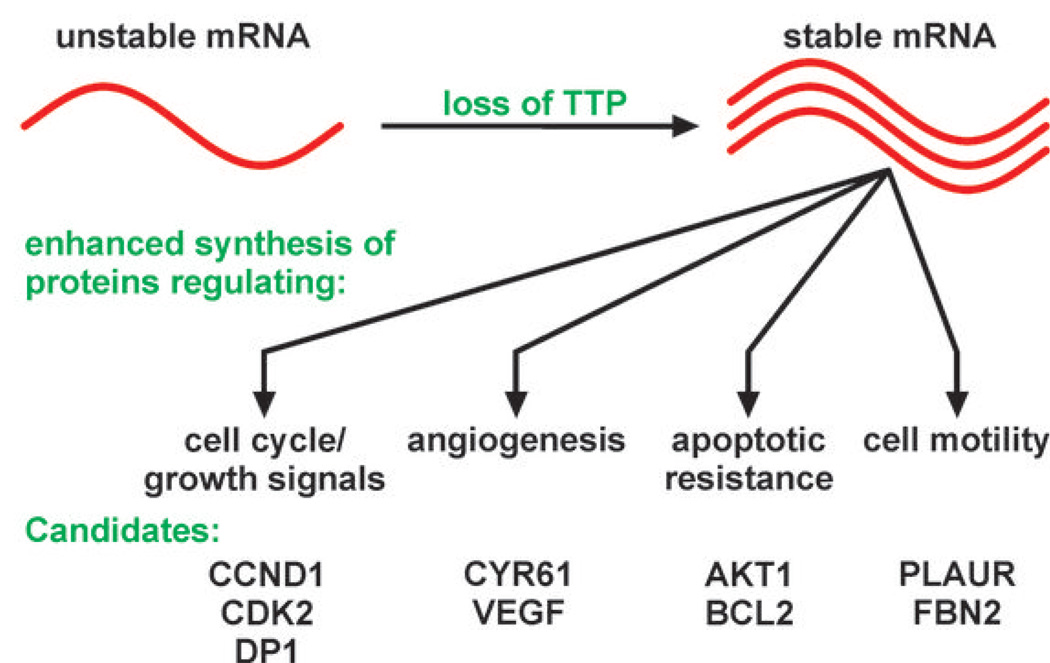
AU-rich element-binding proteins (ARE-BP) regulate the stability and/or translational efficiency of mRNAs containing cognate binding sites. Many targeted transcripts encode factors that control processes such as cell division, apoptosis, and angiogenesis, suggesting that dysregulated ARE-BP expression could dramatically influence oncogenic phenotypes. Using several approaches, we evaluated the expression of four well-characterized ARE-BPs across a variety of human neoplastic syndromes. AUF1, TIA-1, and HuR mRNAs were not systematically dysregulated in cancers; however, tristetraprolin mRNA levels were significantly decreased across many tumor types, including advanced cancers of the breast and prostate. Restoring tristetraprolin expression in an aggressive tumor cell line suppressed three key tumorgenic phenotypes: cell proliferation, resistance to proapoptotic stimuli, and expression of vascular endothelial growth factor mRNA. However, the cellular consequences of tristetraprolin expression varied across different cell models. Analyses of gene array data sets revealed that suppression of tristetraprolin expression is a negative prognostic indicator in breast cancer, because patients with low tumor tristetraprolin mRNA levels were more likely to present increased pathologic tumor grade, vascular endothelial growth factor expression, and mortality from recurrent disease. Collectively, these data establish that tristetraprolin expression is frequently suppressed in human cancers, which in turn can alter tumorigenic phenotypes that influence patient outcomes.
Conflict of interest statement
Potential conflicts of interest: The authors have filed a patent application covering the measurement of TTP expression as an oncological diagnostic and prognostic tool.
Figures
References
-
- Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000;100:57–70. - PubMed
-
- Chen CYA, Shyu AB. AU-rich elements: characterization and importance in mRNA degradation. Trends Biochem Sci. 1995;20:465–470. - PubMed
-
- Guhaniyogi J, Brewer G. Regulation of mRNA stability in mammalian cells. Gene. 2001;265:11–23. - PubMed
-
- Zhang T, Kruys V, Huez G, Gueydan C. AU-rich element-mediated translational control: complexity and multiple activities of trans-acting factors. Biochem Soc Trans. 2002;30:952–958. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
