

A bacterial signal peptide is functional in plants and directs proteins to the secretory pathway
- PMID: 19491306
- PMCID: PMC2724687
- DOI: 10.1093/jxb/erp167
A bacterial signal peptide is functional in plants and directs proteins to the secretory pathway
Abstract
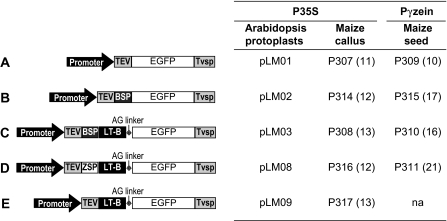
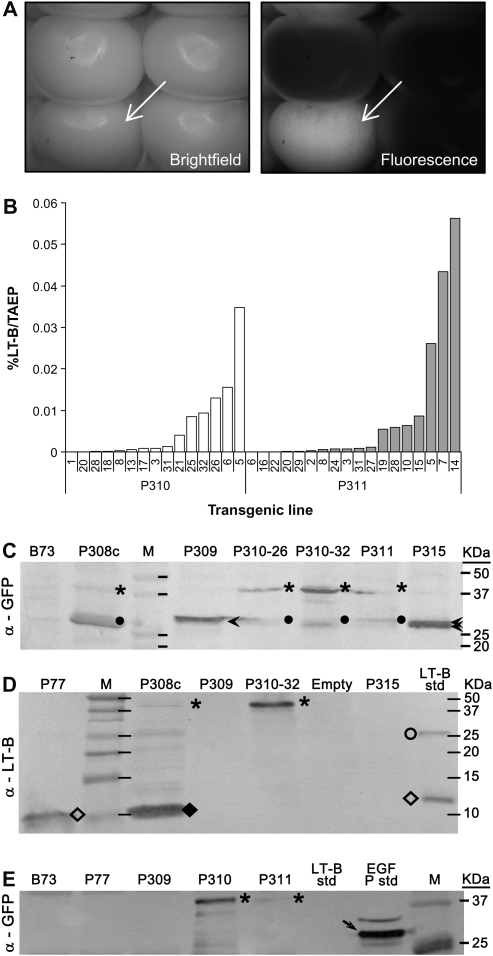
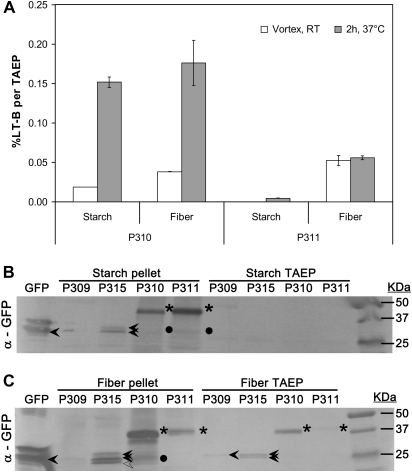
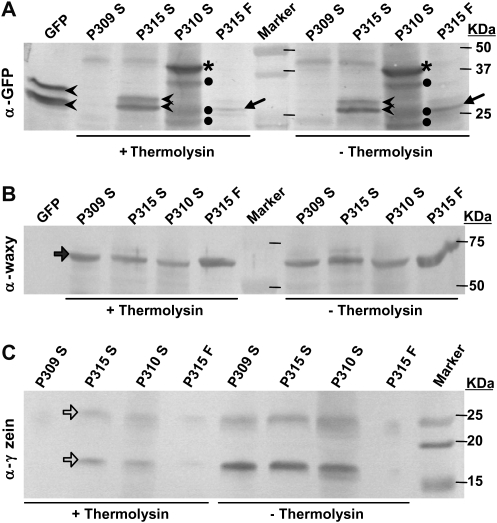
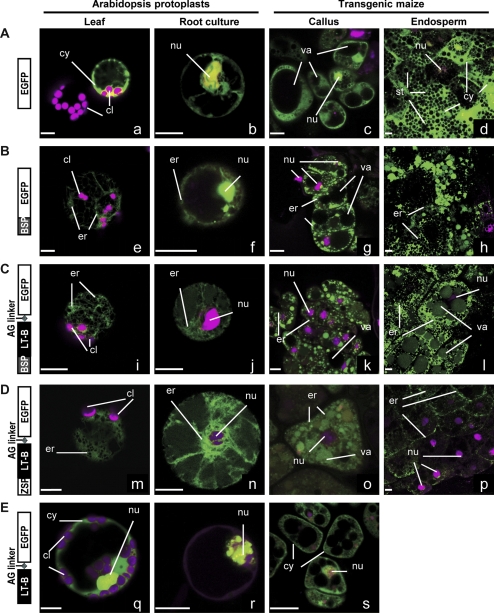
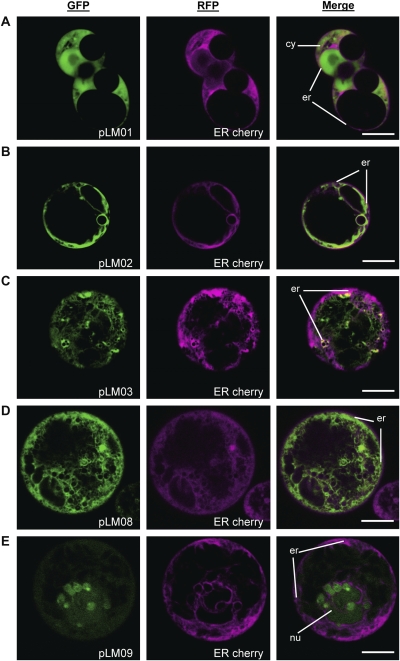
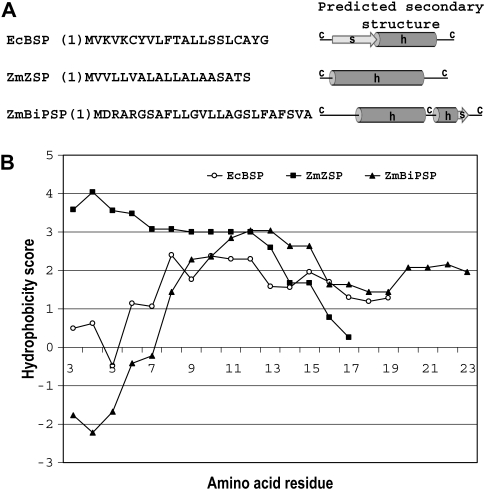
The Escherichia coli heat-labile enterotoxin B subunit (LT-B) has been used as a model antigen for the production of plant-derived high-valued proteins in maize. LT-B with its native signal peptide (BSP) has been shown to accumulate in starch granules of transgenic maize kernels. To elucidate the targeting properties of the bacterial LT-B protein and BSP in plant systems, the subcellular localization of visual marker green fluorescent protein (GFP) fused to LT-B and various combinations of signal peptides was examined in Arabidopsis protoplasts and transgenic maize. Biochemical analysis indicates that the LT-B::GFP fusion proteins can assemble and fold properly retaining both the antigenicity of LT-B and the fluorescing properties of GFP. Maize kernel fractionation revealed that transgenic lines carrying BSP result in recombinant protein association with fibre and starch fractions. Confocal microscopy analysis indicates that the fusion proteins accumulate in the endomembrane system of plant cells in a signal peptide-dependent fashion. This is the first report providing evidence of the ability of a bacterial signal peptide to target proteins to the plant secretory pathway. The results provide important insights for further understanding the heterologous protein trafficking mechanisms and for developing effective strategies in molecular farming.
Figures
Similar articles
-
Characterization of fluorescent chimeras of cholera toxin and Escherichia coli heat-labile enterotoxins produced by use of the twin arginine translocation system.Infect Immun. 2005 Jun;73(6):3627-35. doi: 10.1128/IAI.73.6.3627-3635.2005. Infect Immun. 2005. PMID: 15908392 Free PMC article.
-
Enhanced expression of B-subunit of Escherichia coli heat-labile enterotoxin in tobacco by optimization of coding sequence.Appl Biochem Biotechnol. 2004 Jun;117(3):175-87. doi: 10.1385/abab:117:3:175. Appl Biochem Biotechnol. 2004. PMID: 15304769
-
Production of a fusion protein consisting of the enterotoxigenic Escherichia coli heat-labile toxin B subunit and a tuberculosis antigen in Arabidopsis thaliana.Plant Cell Rep. 2004 Feb;22(7):502-8. doi: 10.1007/s00299-003-0718-2. Epub 2003 Oct 10. Plant Cell Rep. 2004. PMID: 14551732
-
Targeting of cholera toxin and Escherichia coli heat labile toxin in polarized epithelia: role of COOH-terminal KDEL.J Cell Biol. 1995 Nov;131(4):951-62. doi: 10.1083/jcb.131.4.951. J Cell Biol. 1995. PMID: 7490296 Free PMC article.
-
Export and processing analysis of a fusion between the extracellular heat-stable enterotoxin and the periplasmic B subunit of the heat-labile enterotoxin in Escherichia coli.Mol Microbiol. 1990 Feb;4(2):253-64. doi: 10.1111/j.1365-2958.1990.tb00592.x. Mol Microbiol. 1990. PMID: 2187145
Cited by
-
An Interplay between Mitochondrial and ER Targeting of a Bacterial Signal Peptide in Plants.Plants (Basel). 2023 Jan 31;12(3):617. doi: 10.3390/plants12030617. Plants (Basel). 2023. PMID: 36771701 Free PMC article.
-
Ectopic expression of bacterial amylopullulanase enhances bioethanol production from maize grain.Plant Cell Rep. 2013 Sep;32(9):1393-405. doi: 10.1007/s00299-013-1453-y. Epub 2013 May 8. Plant Cell Rep. 2013. PMID: 23652819
-
Recent advances on host plants and expression cassettes' structure and function in plant molecular pharming.BioDrugs. 2014 Apr;28(2):145-59. doi: 10.1007/s40259-013-0062-1. BioDrugs. 2014. PMID: 23959796 Free PMC article. Review.
-
Expression of the cholera toxin B subunit (CT-B) in maize seeds and a combined mucosal treatment against cholera and traveler's diarrhea.Plant Cell Rep. 2012 Mar;31(3):527-37. doi: 10.1007/s00299-011-1146-3. Epub 2011 Sep 22. Plant Cell Rep. 2012. PMID: 21938449
-
Expression of Foreign Genes Demonstrates the Effectiveness of Pollen-Mediated Transformation in Zea mays.Front Plant Sci. 2017 Mar 21;8:383. doi: 10.3389/fpls.2017.00383. eCollection 2017. Front Plant Sci. 2017. PMID: 28377783 Free PMC article.
References
-
- Abramoff MD, Magelhaes PJ, Ram SJ. Image processing with ImageJ. Biophotonics International. 2004;11:36–42.
-
- Asatsuma S, Sawada C, Itoh K, Okito M, Kitajima A, Mitsui T. Involvement of α-amylase I-1 in starch degradation in rice chloroplasts. Plant and Cell Physiology. 2005;46:858–869. - PubMed
-
- Attardi G, Schatz G. Biogenesis of mitochondria. Annual Review of Cell Biology. 1988;4:289–331. - PubMed
-
- Bannai H, Tamada Y, Maruyama O, Nakai K, Miyano S. Extensive feature detection of N-terminal protein sorting signals. Bioinformatics. 2002;18:298–305. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
