

Neutrophil morphology and migration are affected by substrate elasticity
- PMID: 19491394
- PMCID: PMC2727411
- DOI: 10.1182/blood-2008-11-191445
Neutrophil morphology and migration are affected by substrate elasticity
Abstract
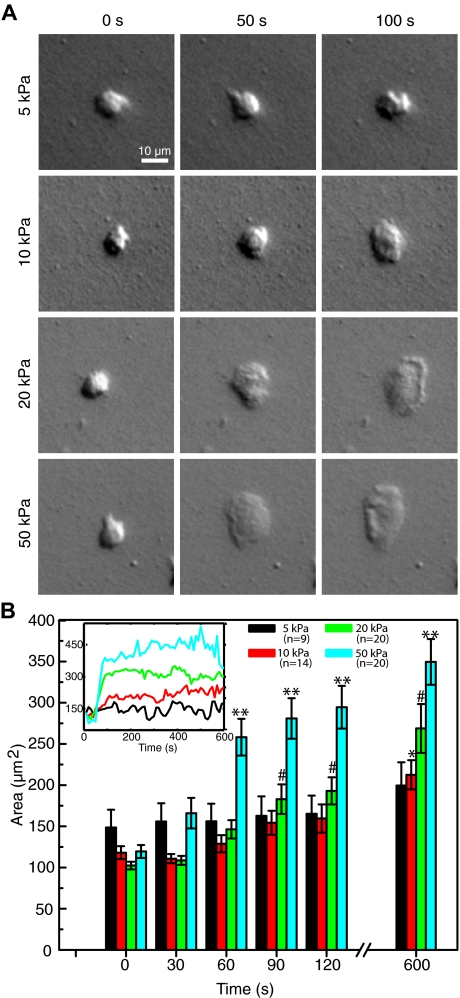
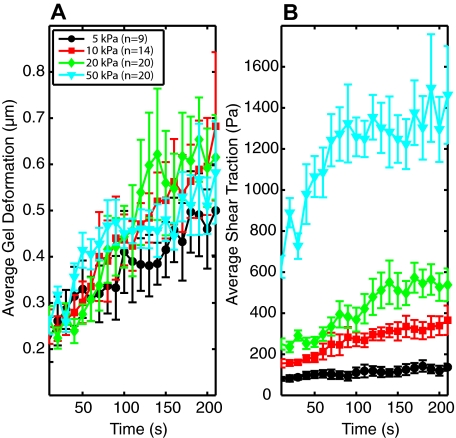
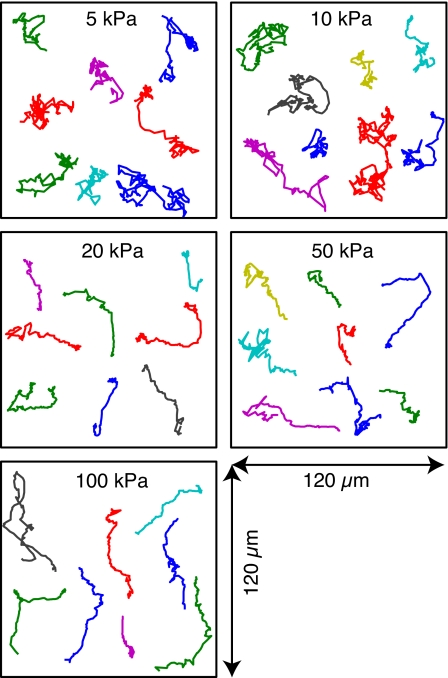
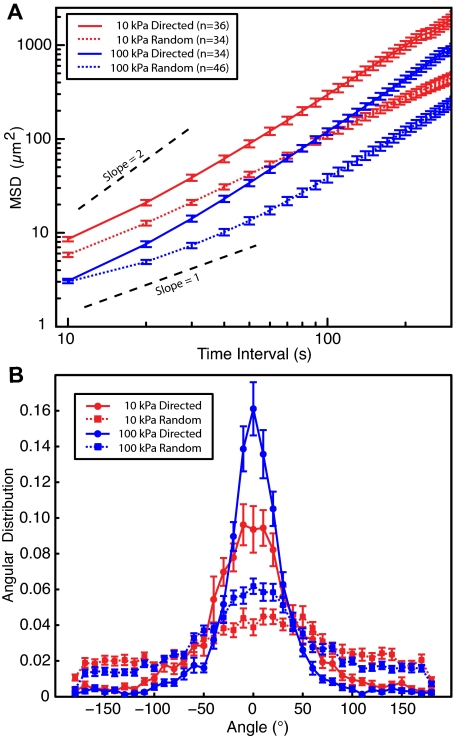
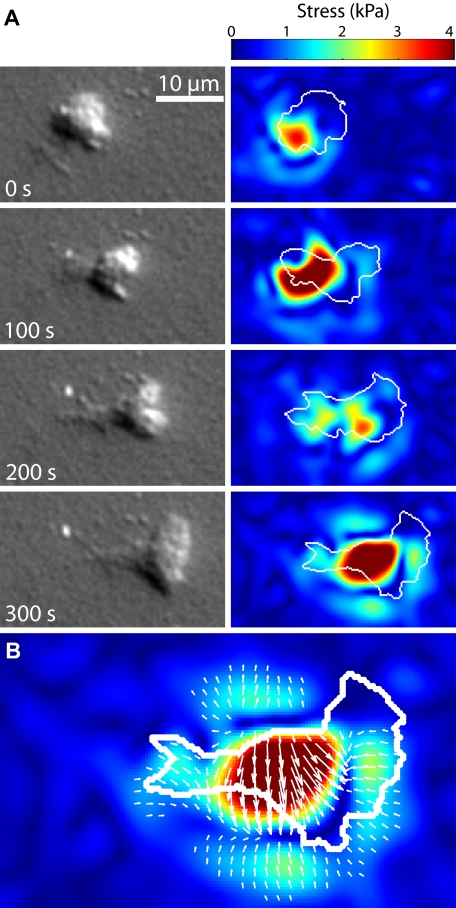
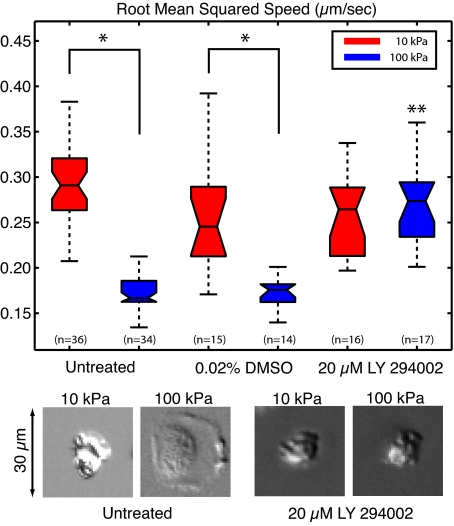
To reach sites of inflammation, neutrophils execute a series of adhesion and migration events that include transmigration through the vascular endothelium and chemotaxis through the vicinal extracellular matrix until contact is made with the point of injury or infection. These in vivo microenvironments differ in their mechanical properties. Using polyacrylamide gels of physiologically relevant elasticity in the range of 5 to 100 kPa and coated with fibronectin, we tested how neutrophil adhesion, spreading, and migration were affected by substrate stiffness. Neutrophils on the softest gels showed only small changes in spread area, whereas on the stiffest gels they showed a 3-fold increase. During adhesion and migration, the magnitudes of the distortions induced in the gel substrate were independent of substrate stiffness, corresponding to the generation of significantly larger traction stresses on the stiffer gels. Cells migrated more slowly but more persistently on stiffer substrates, which resulted in neutrophils moving greater distances over time despite their slower speeds. The largest tractions were localized to the posterior of migrating neutrophils and were independent of substrate stiffness. Finally, the phosphatidylinositol 3-kinase inhibitor LY294002 obviated the ability to sense substrate stiffness, suggesting that phosphatidylinositol 3-kinase plays a mechanistic role in neutrophil mechanosensing.
Figures

Similar articles
-
Effect of Adhesion and Substrate Elasticity on Neutrophil Extracellular Trap Formation.Front Immunol. 2019 Oct 1;10:2320. doi: 10.3389/fimmu.2019.02320. eCollection 2019. Front Immunol. 2019. PMID: 31632402 Free PMC article.
-
Neutrophils display biphasic relationship between migration and substrate stiffness.Cell Motil Cytoskeleton. 2009 Jun;66(6):328-41. doi: 10.1002/cm.20363. Cell Motil Cytoskeleton. 2009. PMID: 19373775
-
Neutrophil adhesion and chemotaxis depend on substrate mechanics.J Phys Condens Matter. 2010 May 19;22(19):194117. doi: 10.1088/0953-8984/22/19/194117. J Phys Condens Matter. 2010. PMID: 20473350 Free PMC article.
-
Physical biology in cancer. 4. Physical cues guide tumor cell adhesion and migration.Am J Physiol Cell Physiol. 2014 Jan 15;306(2):C98-C109. doi: 10.1152/ajpcell.00289.2013. Epub 2013 Oct 16. Am J Physiol Cell Physiol. 2014. PMID: 24133064 Free PMC article. Review.
-
Cell and tissue mechanics in cell migration.Exp Cell Res. 2013 Oct 1;319(16):2418-23. doi: 10.1016/j.yexcr.2013.04.023. Epub 2013 May 9. Exp Cell Res. 2013. PMID: 23664834 Free PMC article. Review.
Cited by
-
Extracellular matrix stiffness-The central cue for skin fibrosis.Front Mol Biosci. 2023 Mar 8;10:1132353. doi: 10.3389/fmolb.2023.1132353. eCollection 2023. Front Mol Biosci. 2023. PMID: 36968277 Free PMC article. Review.
-
Keloid progression: a stiffness gap hypothesis.Int Wound J. 2017 Oct;14(5):764-771. doi: 10.1111/iwj.12693. Epub 2016 Dec 19. Int Wound J. 2017. PMID: 27995750 Free PMC article.
-
Inflammatory Markers for Arterial Stiffness in Cardiovascular Diseases.Front Immunol. 2017 Aug 31;8:1058. doi: 10.3389/fimmu.2017.01058. eCollection 2017. Front Immunol. 2017. PMID: 28912780 Free PMC article. Review.
-
New Tools for Imaging Neutrophils: Work Function Mapping and Element-Specific, Label-Free Imaging of Cellular Structures.Nano Lett. 2021 Jan 13;21(1):222-229. doi: 10.1021/acs.nanolett.0c03554. Epub 2020 Dec 2. Nano Lett. 2021. PMID: 33263404 Free PMC article.
-
LTB4 is a signal-relay molecule during neutrophil chemotaxis.Dev Cell. 2012 May 15;22(5):1079-91. doi: 10.1016/j.devcel.2012.02.003. Epub 2012 Apr 26. Dev Cell. 2012. PMID: 22542839 Free PMC article.
References
-
- Janmey PA, Weitz DA. Dealing with mechanics: mechanisms of force transduction in cells. Trends Biochem Sci. 2004;29:364–370. - PubMed
-
- Discher DE, Janmey P, Wang YL. Tissue cells feel and respond to the stiffness of their substrate. Science. 2005;310:1139–1143. - PubMed
-
- Ali MH, Schumacker PT. Endothelial responses to mechanical stress: where is the mechanosensor? Crit Care Med. 2002;30(suppl):S198–S206. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
