

Monoclonal antibodies recognize distinct conformational epitopes formed by polyglutamine in a mutant huntingtin fragment
- PMID: 19491400
- PMCID: PMC2755888
- DOI: 10.1074/jbc.M109.016923
Monoclonal antibodies recognize distinct conformational epitopes formed by polyglutamine in a mutant huntingtin fragment
Abstract
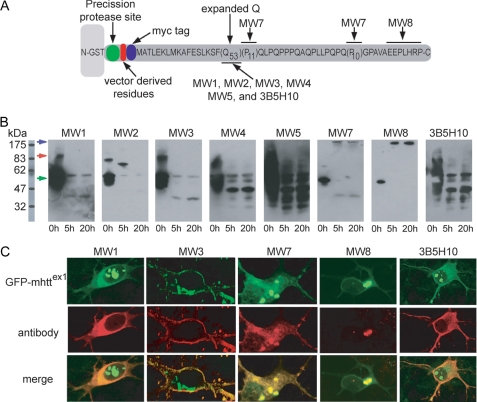
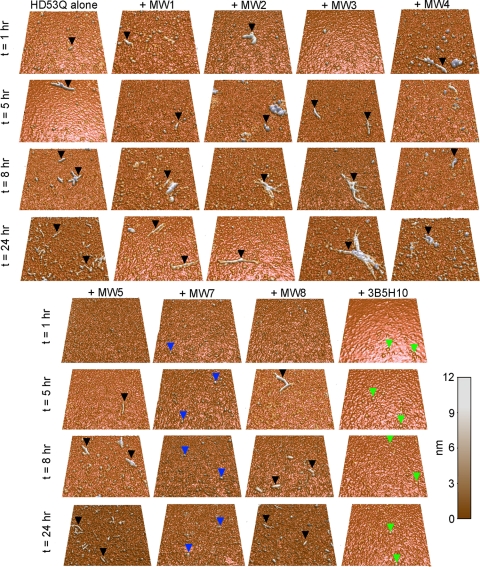
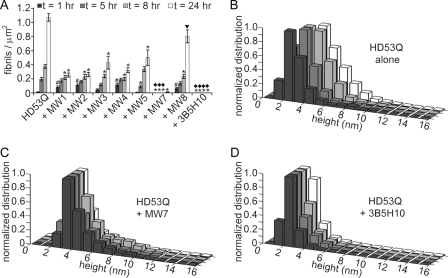
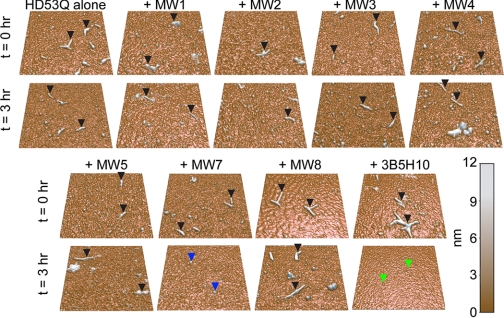
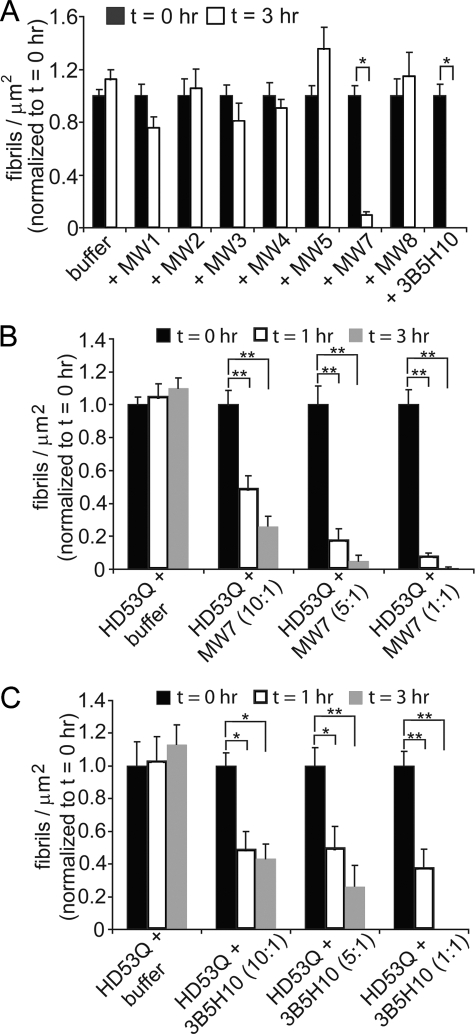
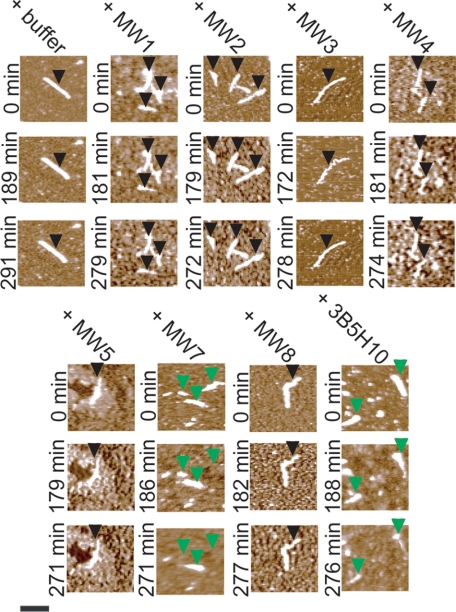
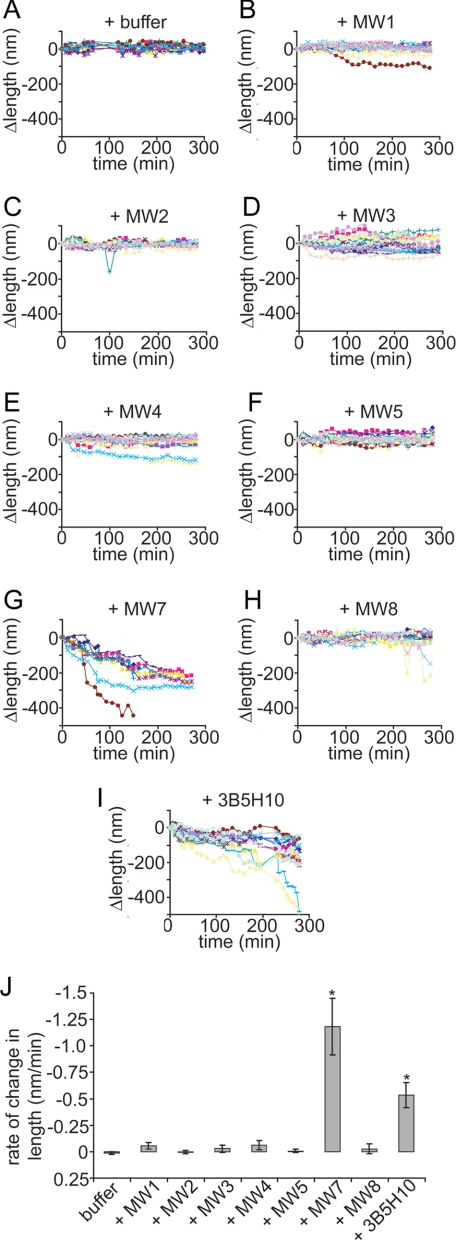
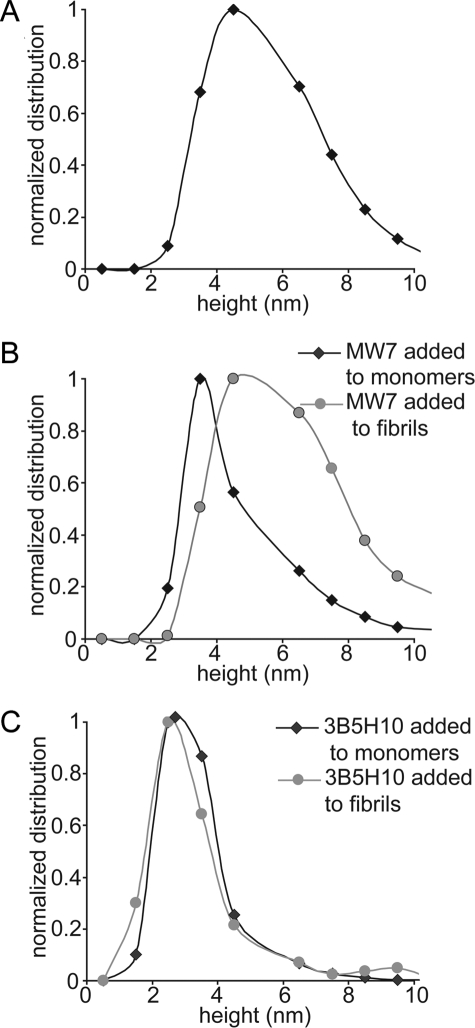
Huntington disease (HD) is a neurodegenerative disorder caused by an expansion of a polyglutamine (polyQ) domain in the N-terminal region of huntingtin (htt). PolyQ expansion above 35-40 results in disease associated with htt aggregation into inclusion bodies. It has been hypothesized that expanded polyQ domains adopt multiple potentially toxic conformations that belong to different aggregation pathways. Here, we used atomic force microscopy to analyze the effect of a panel of anti-htt antibodies (MW1-MW5, MW7, MW8, and 3B5H10) on aggregate formation and the stability of a mutant htt-exon1 fragment. Two antibodies, MW7 (polyproline-specific) and 3B5H10 (polyQ-specific), completely inhibited fibril formation and disaggregated preformed fibrils, whereas other polyQ-specific antibodies had widely varying effects on aggregation. These results suggest that expanded polyQ domains adopt multiple conformations in solution that can be readily distinguished by monoclonal antibodies, which has important implications for understanding the structural basis for polyQ toxicity and the development of intrabody-based therapeutics for HD.
Figures
References
-
- Vonsattel J. P., Myers R. H., Stevens T. J., Ferrante R. J., Bird E. D., Richardson E. P., Jr. (1985) J. Neuropathol. Exp. Neurol. 44, 559–577 - PubMed
-
- Buxbaum J. N. (2003) Trends Biochem. Sci. 28, 585–592 - PubMed
-
- Chiti F., Dobson C. M. (2006) Annu. Rev. Biochem. 75, 333–366 - PubMed
-
- Ross C. A., Poirier M. A. (2004) Nat. Med. 10, S10–S17 - PubMed
-
- Tobin A. J., Signer E. R. (2000) Trends Cell Biol. 10, 531–536 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01NS054753/NS/NINDS NIH HHS/United States
- P01AG022074/AG/NIA NIH HHS/United States
- R01 NS055298/NS/NINDS NIH HHS/United States
- R01NS039074/NS/NINDS NIH HHS/United States
- P01 AG022074/AG/NIA NIH HHS/United States
- R01 NS054753/NS/NINDS NIH HHS/United States
- R01NS047237/NS/NINDS NIH HHS/United States
- R01NS045091/NS/NINDS NIH HHS/United States
- R01NS055298/NS/NINDS NIH HHS/United States
- R01 NS039074/NS/NINDS NIH HHS/United States
- R01 NS047237/NS/NINDS NIH HHS/United States
- R01 NS045091/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
