

Cellular mechanisms of membrane protein folding
- PMID: 19491932
- PMCID: PMC2814870
- DOI: 10.1038/nsmb.1600
Cellular mechanisms of membrane protein folding
Abstract
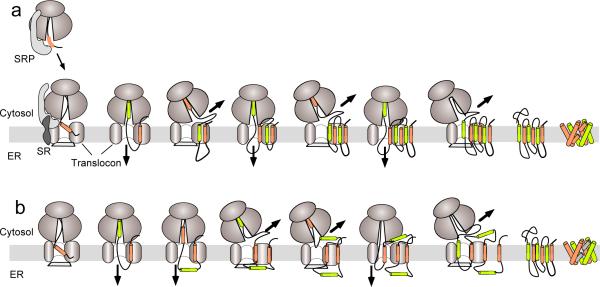
The membrane protein-folding problem can be articulated by two central questions. How is protein topology established by selective peptide transport to opposite sides of the cellular membrane? And how are transmembrane segments inserted, integrated and folded within the lipid bilayer? In eukaryotes, this process usually takes place in the endoplasmic reticulum, coincident with protein synthesis, and is facilitated by the translating ribosome and the Sec61 translocon complex (RTC). At its core, the RTC forms a dynamic pathway through which the elongating nascent polypeptide moves as it is delivered into the cytosolic, lumenal and lipid compartments. This Perspective will focus on emerging evidence that the RTC functions as a protein-folding machine that restricts conformational space by establishing transmembrane topology and yet provides a permissive environment that enables nascent transmembrane domains to efficiently progress down their folding energy landscape.
Figures


References
-
- Popot J, Engelman D. Helical membrane protein folding, stability and evolution. Ann. Rev. Biochem. 2000;69:881–922. - PubMed
-
- Bowie J. Solving the membrane protein folding problem. Nature. 2005;438:581–589. - PubMed
-
- Johnson A, van Waes M. The Translocon: A dynamic gateway at the ER membrane. Ann. Rev. Cell Dev. Biol. 1999;15:799–842. - PubMed
-
- Rapoport T, Goder V, Heinrich S, Matlack K. Membrane-protein integration and the role of the translocation channel. Trends Cell Biol. 2004;14:568–575. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
