

Evidence for two retinoid cycles in the cone-dominated chicken eye
- PMID: 19492794
- PMCID: PMC2733357
- DOI: 10.1021/bi9002937
Evidence for two retinoid cycles in the cone-dominated chicken eye
Abstract
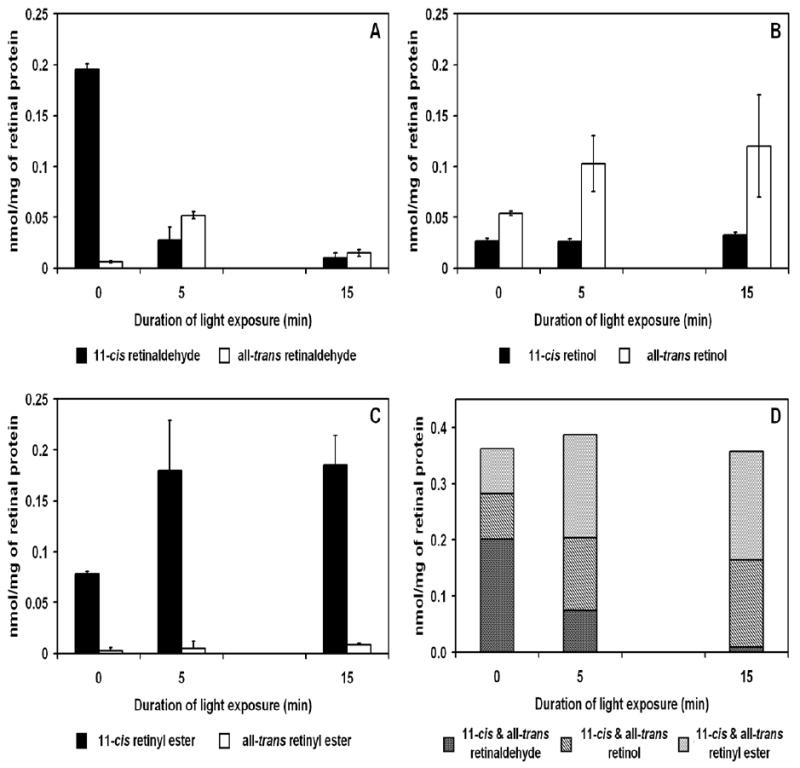
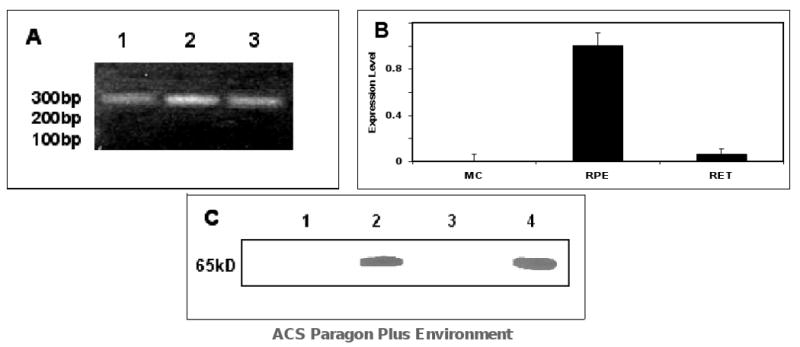
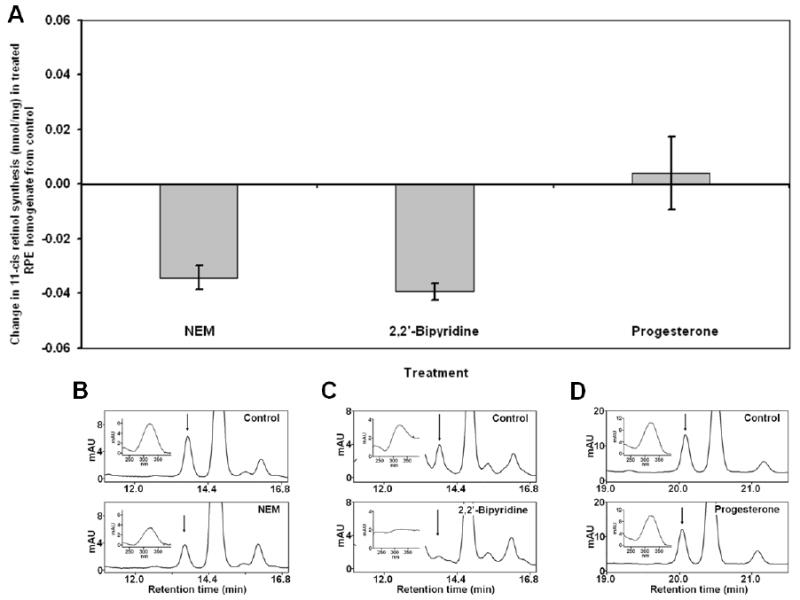
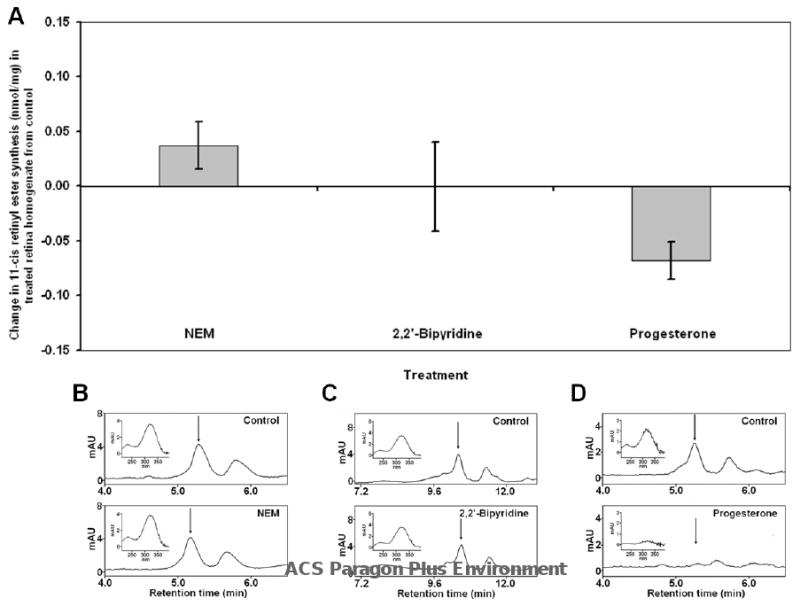
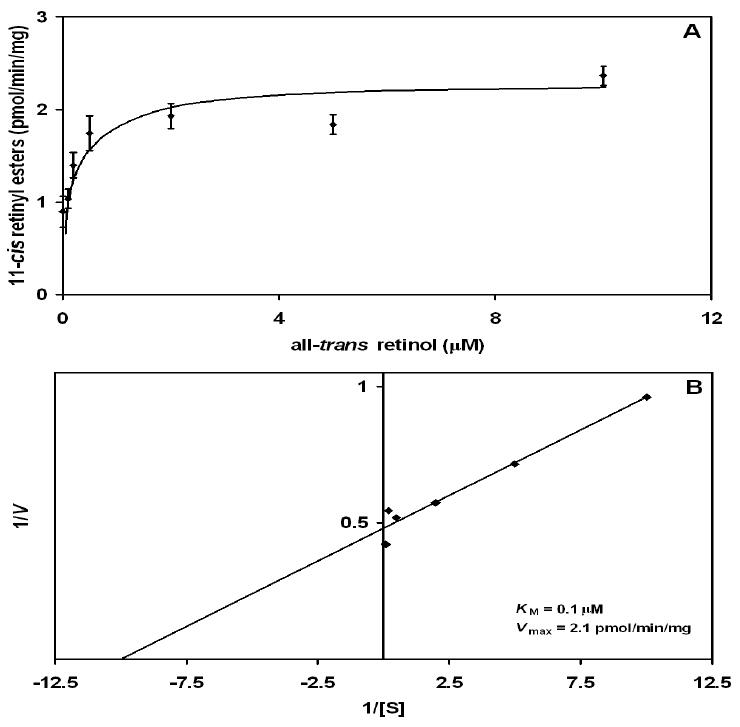
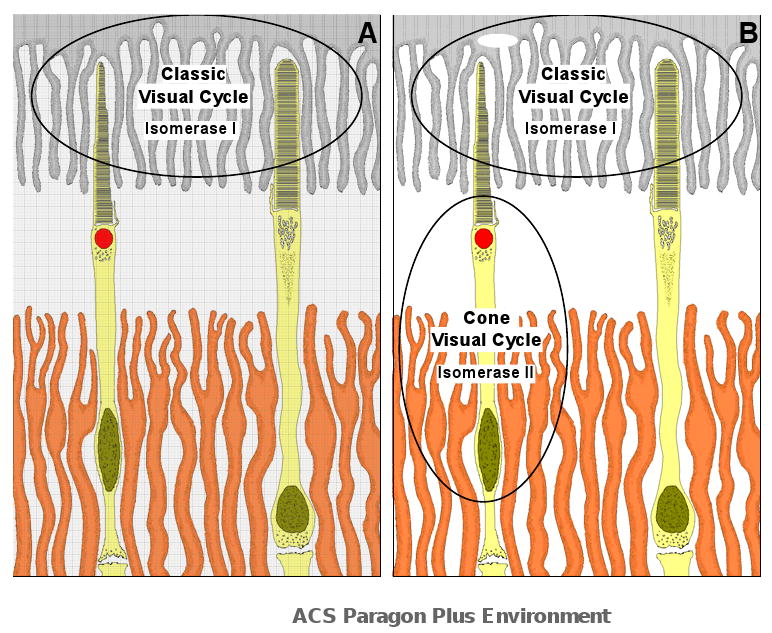
In the classic retinoid cycle, 11-cis retinol is synthesized in the retinal pigment epithelium (RPE) by two enzymes: Isomerase I (RPE65) and lecithin:retinol acyltransferase (LRAT). The purpose of this study is to provide experimental evidence for two active isomerases in the cone-dominated chicken eye: an LRAT-dependent Isomerase I in the RPE and an ARAT (acyl CoA:retinol acyltransferase)-dependent isomerase (Isomerase II) in the retina. First, we show that whole chicken retina in vitro, removed from the RPE/choroid and sclera, produces 11-cis retinoids upon light exposure, indicating the existence of RPE-independent isomerase (Isomerase II) activity in the retina. Reverse transcriptase polymerase chain reaction studies show high levels of RPE65 expression in the RPE, low levels in the retina, and none in primary Muller cell cultures, indicating the presence of Isomerase I in the RPE and a minimal amount in the retina. Activities of the RPE and retina isomerases were then measured by enzyme assays with specific enzyme inhibitors. 2,2'-Bipyridine, a known Isomerase I inhibitor, and N-ethylmaleimide (NEM), a known LRAT inhibitor, significantly reduced Isomerase I activity but not Isomerase II activity. Progesterone, a known ARAT inhibitor, completely blocked Isomerase II activity but not Isomerase I activity. Thus, this study reports novel results for distinguishing the biochemical properties of Isomerase I from those of Isomerase II, as well a difference in their locations in the chicken eye. On the basis of these differences, the cone-dominated chicken eye must contain two retinoid cycles: a classic visual cycle for retinoid exchange between the RPE and the retina supported by Isomerase I in the RPE and an additional visual cycle for retinoid processing in the retina supported by Isomerase II.
Figures
References
-
- Crouch RK, Chader GJ, Wiggert B, Pepperberg DR. Retinoids and the visual process. Photochem Photobiol. 1996;64(4):613–621. - PubMed
-
- Saari JC. Biochemistry of visual pigment regeneration: the Friedenwald lecture. Invest Ophthalmol Vis Sci. 2000;41(2):337–348. - PubMed
-
- McBee JK, Palczewski K, Baehr W, Pepperberg DR. Confronting complexity: the interlink of phototransduction and retinoid metabolism in the vertebrate retina. Prog Retin Eye Res. 2001;20(4):469–529. - PubMed
-
- Rando RR. The biochemistry of the visual cycle. Chem Rev. 2001;101(7):1881–1896. - PubMed
-
- Lamb TD, Pugh EN., Jr Dark adaptation and the retinoid cycle of vision. Prog Retin Eye Res. 2004;23(3):307–380. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
