

Sphingosine-1-phosphate receptors mediate neuromodulatory functions in the CNS
- PMID: 19493165
- PMCID: PMC2754148
- DOI: 10.1111/j.1471-4159.2009.06202.x
Sphingosine-1-phosphate receptors mediate neuromodulatory functions in the CNS
Abstract
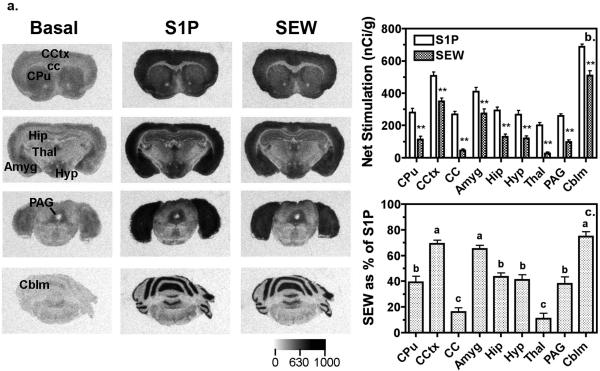
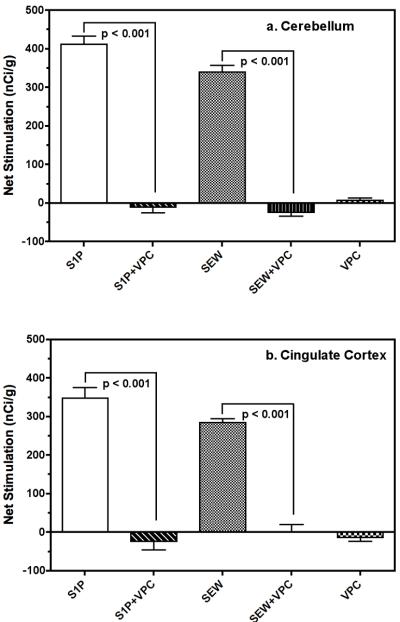
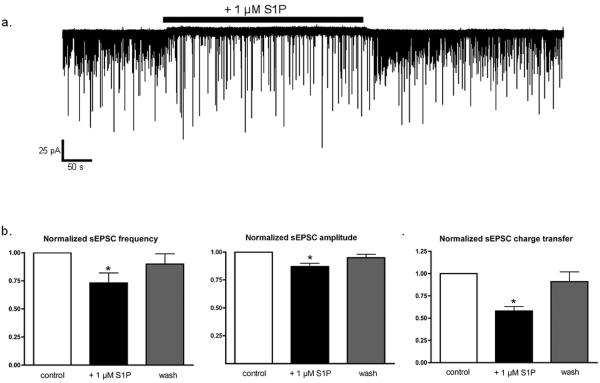
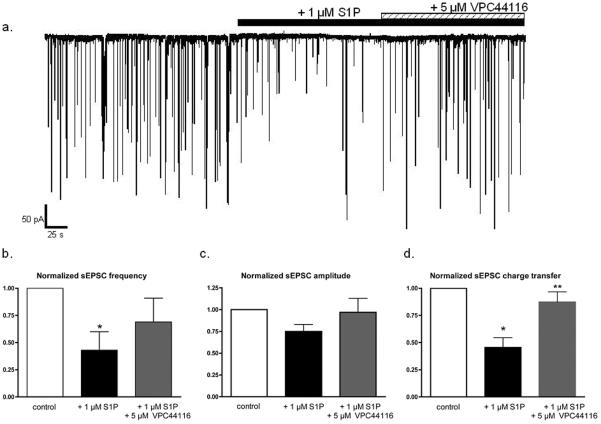
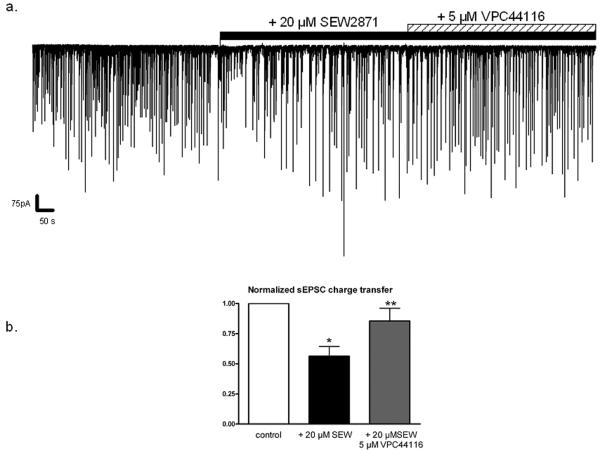
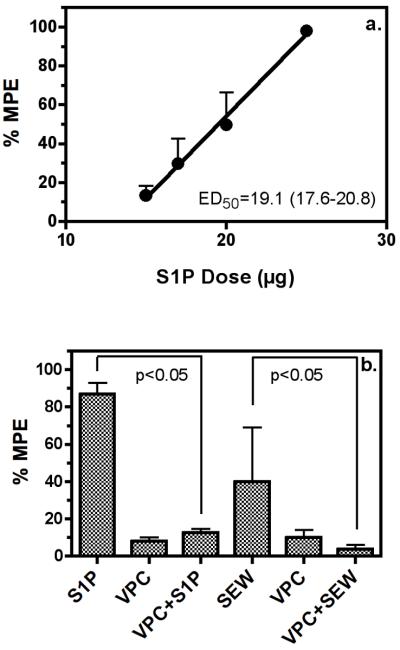
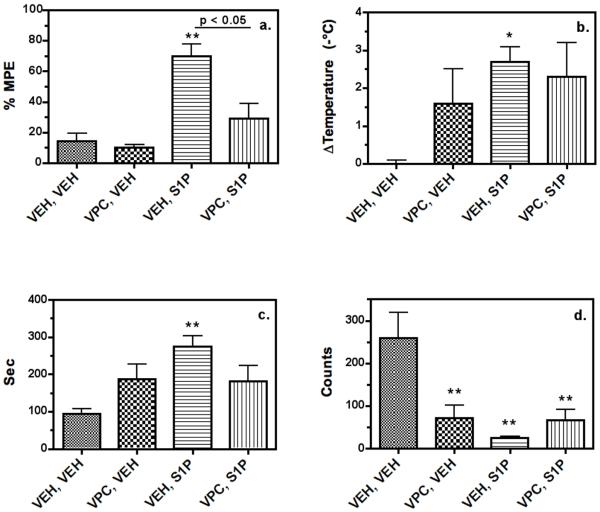
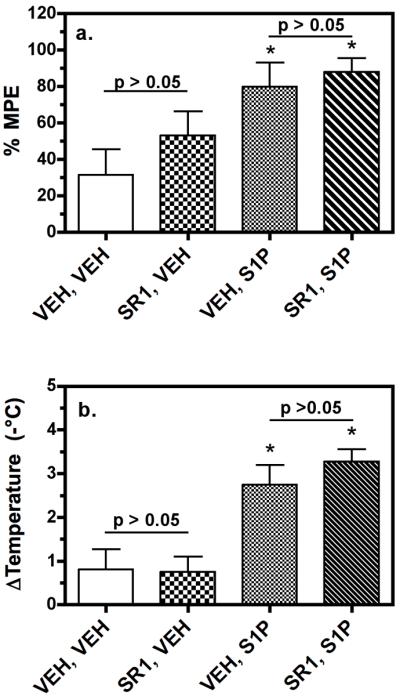
Sphingosine-1-phosphate (S1P) is a ubiquitous, lipophilic cellular mediator that acts in part by activation of G-protein-coupled receptor. Modulation of S1P signaling is an emerging pharmacotherapeutic target for immunomodulatory drugs. Although multiple S1P receptor types exist in the CNS, little is known about their function. Here, we report that S1P stimulated G-protein activity in the CNS, and results from [(35)S]GTPgammaS autoradiography using the S1P(1)-selective agonist SEW2871 and the S1P(1/3)-selective antagonist VPC44116 show that in several regions a majority of this activity is mediated by S1P(1) receptors. S1P receptor activation inhibited glutamatergic neurotransmission as determined by electrophysiological recordings in cortical neurons in vitro, and this effect was mimicked by SEW2871 and inhibited by VPC44116. Moreover, central administration of S1P produced in vivo effects resembling the actions of cannabinoids, including thermal antinociception, hypothermia, catalepsy and hypolocomotion, but these actions were independent of CB(1) receptors. At least one of the central effects of S1P, thermal antinociception, is also at least partly S1P(1) receptor mediated because it was produced by SEW2871 and attenuated by VPC44116. These results indicate that CNS S1P receptors are part of a physiologically relevant and widespread neuromodulatory system, and that the S1P(1) receptor contributes to S1P-mediated antinociception.
Figures
References
-
- Beer MS, Stanton JA, Salim K, Rigby M, Heavens RP, Smith D, McAllister G. EDG receptors as a therapeutic target in the nervous system. Ann. N. Y. Acad. Sci. 2000;905:118–131. - PubMed
-
- Bliss CI. Statistics in Biology. McGraw-Hill; New York: 1967.
-
- Brinkmann V. Sphingosine 1-phosphate receptors in health and disease: Mechanistic insights from gene deletion studies and reverse pharmacology. Pharmacol. Ther. 2007;115:84–105. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
