

Genome-wide analysis of plastid gene expression in potato leaf chloroplasts and tuber amyloplasts: transcriptional and posttranscriptional control
- PMID: 19493969
- PMCID: PMC2719133
- DOI: 10.1104/pp.109.140483
Genome-wide analysis of plastid gene expression in potato leaf chloroplasts and tuber amyloplasts: transcriptional and posttranscriptional control
Abstract
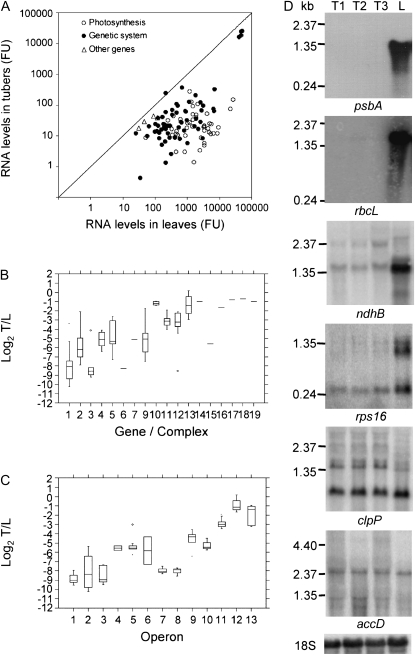
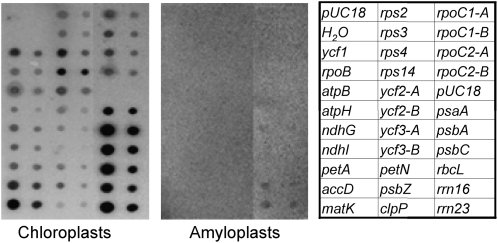
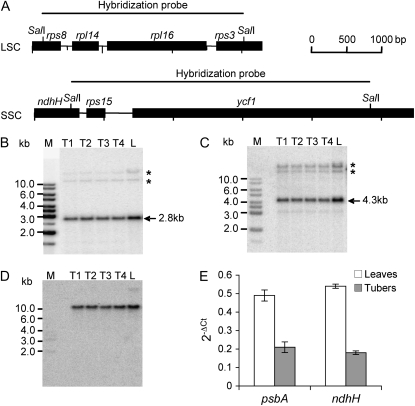
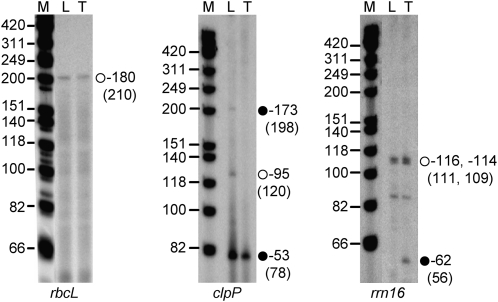
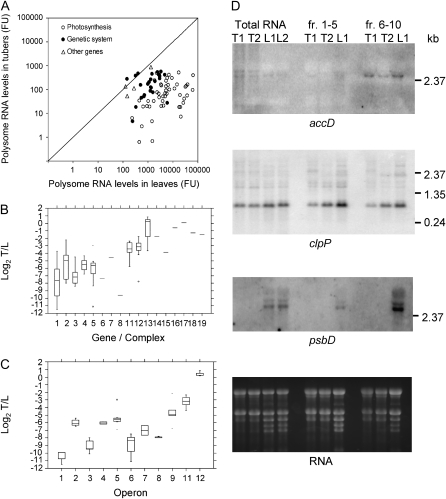
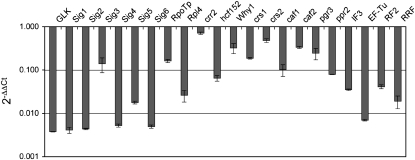
Gene expression in nongreen plastids is largely uncharacterized. To compare gene expression in potato (Solanum tuberosum) tuber amyloplasts and leaf chloroplasts, amounts of transcripts of all plastid genes were determined by hybridization to plastome arrays. Except for a few genes, transcript accumulation was much lower in tubers compared with leaves. Transcripts of photosynthesis-related genes showed a greater reduction in tubers compared with leaves than transcripts of genes for the genetic system. Plastid genome copy number in tubers was 2- to 3-fold lower than in leaves and thus cannot account for the observed reduction of transcript accumulation in amyloplasts. Both the plastid-encoded and the nucleus-encoded RNA polymerases were active in potato amyloplasts. Transcription initiation sites were identical in chloroplasts and amyloplasts, although some differences in promoter utilization between the two organelles were evident. For some intron-containing genes, RNA splicing was less efficient in tubers than in leaves. Furthermore, tissue-specific differences in editing of ndh transcripts were detected. Hybridization of the plastome arrays with RNA extracted from polysomes indicated that, in tubers, ribosome association of transcripts was generally low. Nevertheless, some mRNAs, such as the transcript of the fatty acid biosynthesis gene accD, displayed relatively high ribosome association. Selected nuclear genes involved in plastid gene expression were generally significantly less expressed in tubers than in leaves. Hence, compared with leaf chloroplasts, gene expression in tuber amyloplasts is much lower, with control occurring at the transcriptional, posttranscriptional, and translational levels. Candidate regulatory sequences that potentially can improve plastid (trans)gene expression in amyloplasts have been identified.
Figures
References
-
- Adam Z (2007) Protein stability and degradation in plastids. Top Curr Genet 19 315–338
-
- Aguettaz P, Seyer P, Pesey H, Lescure AM (1987) Relations between the plastid gene dosage and the levels of 16S rRNA and rbcL gene transcripts during amyloplast to chloroplast change in mixotrophic spinach cell suspensions. Plant Mol Biol 8 169–177 - PubMed
-
- Allison LA (2000) The role of sigma factors in plastid transcription. Biochimie 82 537–548 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
