

Distinct roles of two zebrafish AHR repressors (AHRRa and AHRRb) in embryonic development and regulating the response to 2,3,7,8-tetrachlorodibenzo-p-dioxin
- PMID: 19494032
- PMCID: PMC2708601
- DOI: 10.1093/toxsci/kfp116
Distinct roles of two zebrafish AHR repressors (AHRRa and AHRRb) in embryonic development and regulating the response to 2,3,7,8-tetrachlorodibenzo-p-dioxin
Abstract
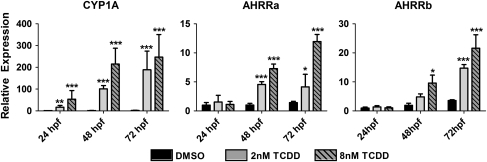
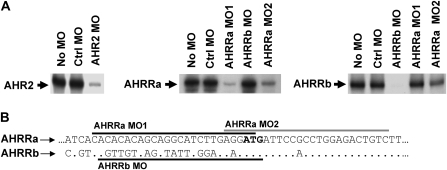
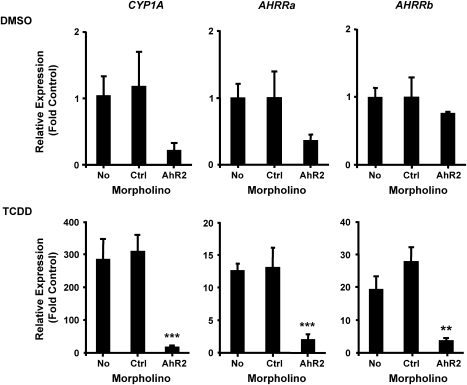
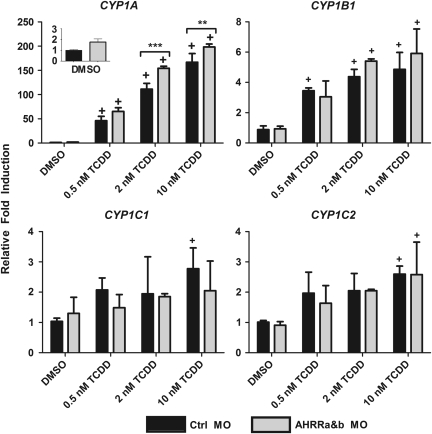
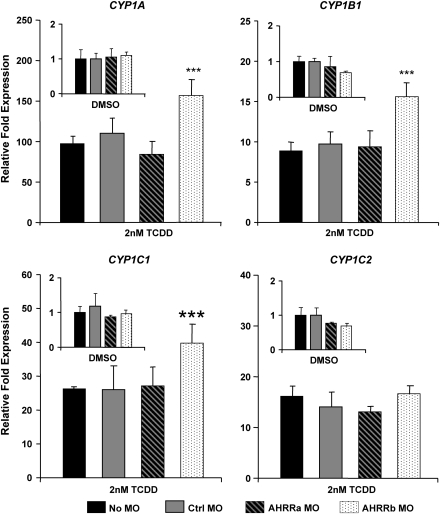
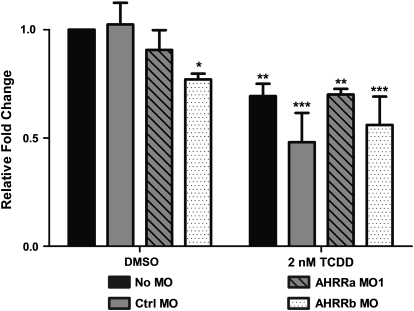
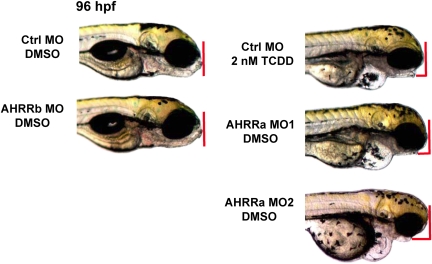
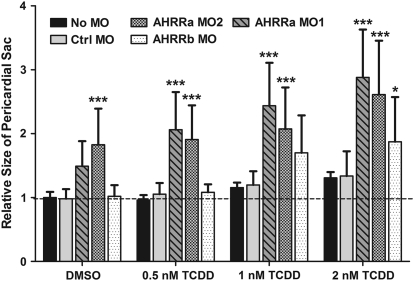
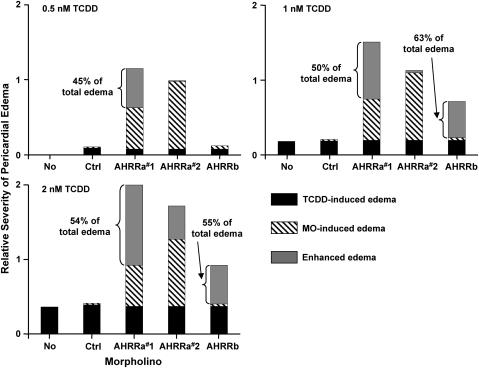
The aryl hydrocarbon receptor (AHR) repressor (AHRR), an AHR-related basic helix-loop-helix/Per-AHR nuclear translocator-Sim protein, is regulated by an AHR-dependent mechanism and acts as a transcriptional repressor of AHR function. Resulting from a teleost-specific genome duplication, zebrafish have two AHRR genes (AHRRa and AHRRb), but their functions in vivo are not well understood. We used antisense morpholino oligonucleotides (MOs) in zebrafish embryos and a zebrafish liver cell line (ZF-L) to characterize the interaction of AHRRs and AHRs in normal embryonic development, AHR signaling, and 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) toxicity. Zebrafish embryos exposed to TCDD (2 and 8nM) during early development showed strong induction of CYP1A, AHRRa, and AHRRb at 48 and 72 hours post-fertilization (hpf). An MO targeting AHR2 inhibited TCDD-induced expression of CYP1A, AHRRa, and AHRRb by 84-95% in 48 hpf embryos, demonstrating a primary role for AHR2 in mediating AHRR induction. Dual MO knockdown of both AHRRs in ZF-L cells enhanced TCDD induction of CYP1A, but not other CYP1 genes. In embryos, dual knockdown of AHRRs, or knockdown of AHRRb alone, enhanced the induction of CYP1A, CYP1B1, and CYP1C1 by TCDD and decreased the constitutive expression of Sox9b. In contrast, knockdown of AHRRa did not affect Sox9b expression or CYP1 inducibility. Embryos microinjected with each of two different MOs targeting AHRRa and exposed to dimethyl sulfoxide (DMSO) displayed developmental phenotypes resembling those typical of TCDD-exposed embryos (pericardial edema and lower jaw malformations). In contrast, no developmental phenotypes were observed in DMSO-exposed AHRRb morphants. These data demonstrate distinct roles of AHRRa and AHRRb in regulating AHR signaling in vivo and suggest that they have undergone subfunction partitioning since the teleost-specific genome duplication.
Figures
Similar articles
-
Knockdown of a zebrafish aryl hydrocarbon receptor repressor (AHRRa) affects expression of genes related to photoreceptor development and hematopoiesis.Toxicol Sci. 2014 Jun;139(2):381-95. doi: 10.1093/toxsci/kfu052. Epub 2014 Mar 27. Toxicol Sci. 2014. PMID: 24675095 Free PMC article.
-
Role of AHR2 in the expression of novel cytochrome P450 1 family genes, cell cycle genes, and morphological defects in developing zebra fish exposed to 3,3',4,4',5-pentachlorobiphenyl or 2,3,7,8-tetrachlorodibenzo-p-dioxin.Toxicol Sci. 2007 Nov;100(1):180-93. doi: 10.1093/toxsci/kfm207. Epub 2007 Aug 8. Toxicol Sci. 2007. PMID: 17686920
-
Zebrafish cardiotoxicity: the effects of CYP1A inhibition and AHR2 knockdown following exposure to weak aryl hydrocarbon receptor agonists.Environ Sci Pollut Res Int. 2015 Jun;22(11):8329-38. doi: 10.1007/s11356-014-3969-2. Epub 2014 Dec 23. Environ Sci Pollut Res Int. 2015. PMID: 25532870 Free PMC article.
-
Understanding dioxin developmental toxicity using the zebrafish model.Birth Defects Res A Clin Mol Teratol. 2006 Jan;76(1):7-18. doi: 10.1002/bdra.20216. Birth Defects Res A Clin Mol Teratol. 2006. PMID: 16333842 Review.
-
Mechanisms of Developmental Toxicity of Dioxins and Related Compounds.Int J Mol Sci. 2019 Jan 31;20(3):617. doi: 10.3390/ijms20030617. Int J Mol Sci. 2019. PMID: 30708991 Free PMC article. Review.
Cited by
-
Predicting the sensitivity of fishes to dioxin-like compounds: possible role of the aryl hydrocarbon receptor (AhR) ligand binding domain.Environ Sci Pollut Res Int. 2013 Mar;20(3):1219-24. doi: 10.1007/s11356-012-1203-7. Epub 2012 Oct 11. Environ Sci Pollut Res Int. 2013. PMID: 23054770 Review.
-
Developmental Regulation of Nuclear Factor Erythroid-2 Related Factors (nrfs) by AHR1b in Zebrafish (Danio rerio).Toxicol Sci. 2019 Feb 1;167(2):536-545. doi: 10.1093/toxsci/kfy257. Toxicol Sci. 2019. PMID: 30321412 Free PMC article.
-
Knockdown of a zebrafish aryl hydrocarbon receptor repressor (AHRRa) affects expression of genes related to photoreceptor development and hematopoiesis.Toxicol Sci. 2014 Jun;139(2):381-95. doi: 10.1093/toxsci/kfu052. Epub 2014 Mar 27. Toxicol Sci. 2014. PMID: 24675095 Free PMC article.
-
Developmental expression of the Nfe2-related factor (Nrf) transcription factor family in the zebrafish, Danio rerio.PLoS One. 2013 Oct 24;8(10):e79574. doi: 10.1371/journal.pone.0079574. eCollection 2013. PLoS One. 2013. PMID: 24298298 Free PMC article.
-
Gene co-expression network analysis in zebrafish reveals chemical class specific modules.BMC Genomics. 2021 Sep 13;22(1):658. doi: 10.1186/s12864-021-07940-4. BMC Genomics. 2021. PMID: 34517816 Free PMC article.
References
-
- Amores A, Force A, Yan Y-L, Joly L, Amemiya C, Fritz A, Ho RK, Langeland J, Prince V, Wang Y-L, et al. Zebrafish hox clusters and vertebrate genome evolution. Science. 1998;282:1711–1714. - PubMed
-
- Andreasen EA, Hahn ME, Heideman W, Peterson RE, Tanguay RL. The zebrafish (Danio rerio) aryl hydrocarbon receptor type 1 is a novel vertebrate receptor. Mol. Pharmacol. 2002;62:234–249. - PubMed
-
- Andreasen EA, Mathew LK, Lohr CV, Hasson R, Tanguay RL. Aryl hydrocarbon receptor activation impairs extracellular matrix remodeling during zebrafish fin regeneration. Toxicol. Sci. 2007;95:215–226. - PubMed
-
- Andreasen EA, Mathew LK, Tanguay RL. Regenerative growth is impacted by TCDD: Gene expression analysis reveals extracellular matrix modulation. Toxicol. Sci. 2006;92:254–269. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
