

Enhancing CD8 T-cell memory by modulating fatty acid metabolism
- PMID: 19494812
- PMCID: PMC2803086
- DOI: 10.1038/nature08097
Enhancing CD8 T-cell memory by modulating fatty acid metabolism
Abstract
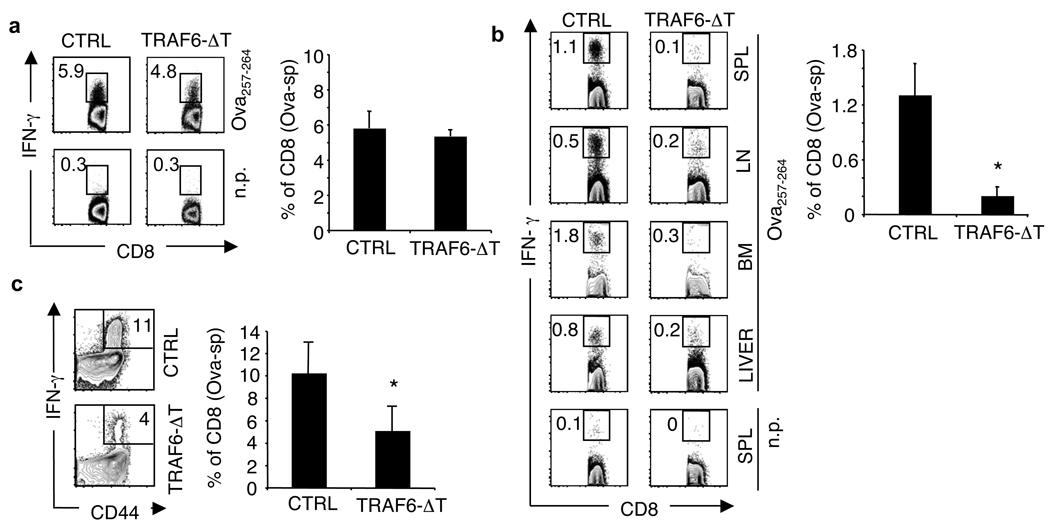
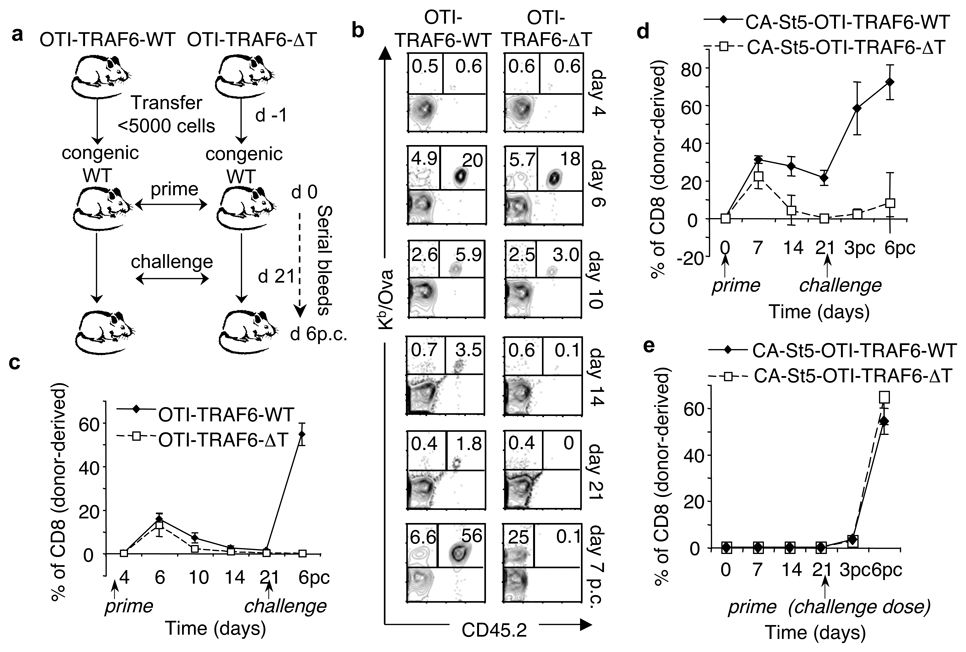
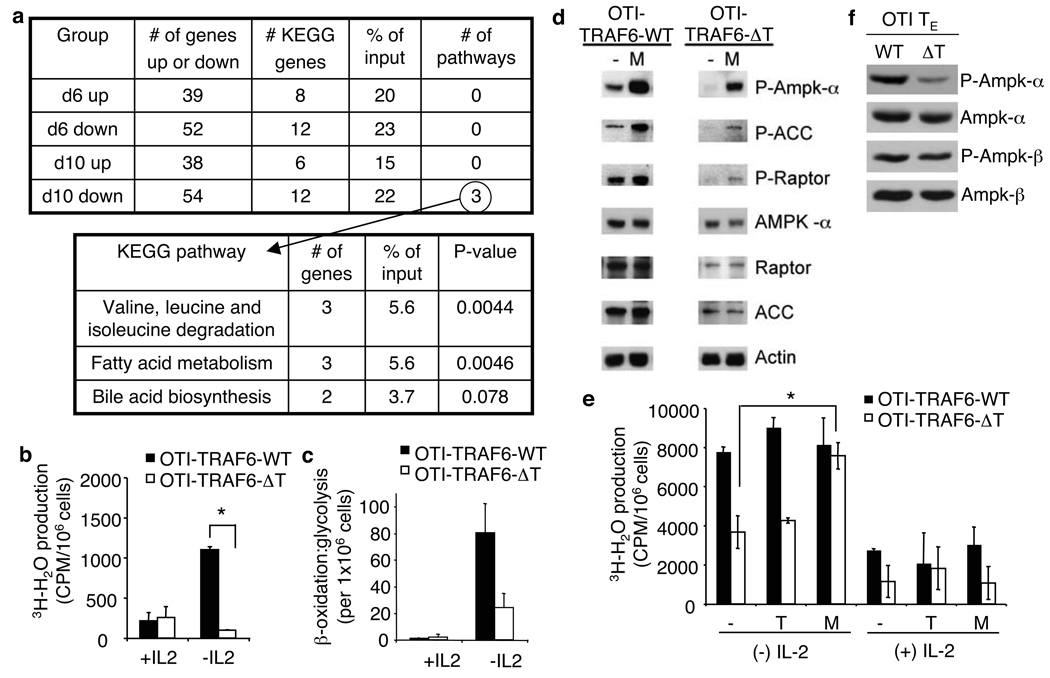
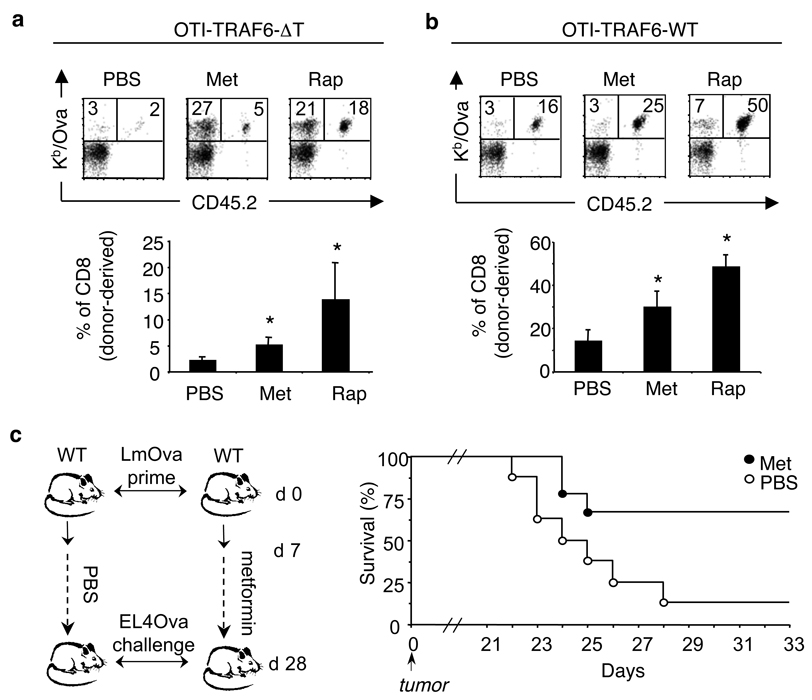
CD8 T cells, which have a crucial role in immunity to infection and cancer, are maintained in constant numbers, but on antigen stimulation undergo a developmental program characterized by distinct phases encompassing the expansion and then contraction of antigen-specific effector (T(E)) populations, followed by the persistence of long-lived memory (T(M)) cells. Although this predictable pattern of CD8 T-cell responses is well established, the underlying cellular mechanisms regulating the transition to T(M) cells remain undefined. Here we show that tumour necrosis factor (TNF) receptor-associated factor 6 (TRAF6), an adaptor protein in the TNF-receptor and interleukin-1R/Toll-like receptor superfamily, regulates CD8 T(M)-cell development after infection by modulating fatty acid metabolism. We show that mice with a T-cell-specific deletion of TRAF6 mount robust CD8 T(E)-cell responses, but have a profound defect in their ability to generate T(M) cells that is characterized by the disappearance of antigen-specific cells in the weeks after primary immunization. Microarray analyses revealed that TRAF6-deficient CD8 T cells exhibit altered expression of genes that regulate fatty acid metabolism. Consistent with this, activated CD8 T cells lacking TRAF6 display defective AMP-activated kinase activation and mitochondrial fatty acid oxidation (FAO) in response to growth factor withdrawal. Administration of the anti-diabetic drug metformin restored FAO and CD8 T(M)-cell generation in the absence of TRAF6. This treatment also increased CD8 T(M) cells in wild-type mice, and consequently was able to considerably improve the efficacy of an experimental anti-cancer vaccine.
Figures
Comment in
-
Immunology: A metabolic switch to memory.Nature. 2009 Jul 2;460(7251):41-2. doi: 10.1038/460041a. Nature. 2009. PMID: 19571872 No abstract available.
References
-
- Harty JT, Badovinac VP. Shaping and reshaping CD8+ T-cell memory. Nat Rev Immunol. 2008;8:107–119. - PubMed
-
- Prlic M, Williams MA, Bevan MJ. Requirements for CD8 T-cell priming, memory generation and maintenance. Curr Opin Immunol. 2007;19:315–319. - PubMed
-
- Lanzavecchia A, Sallusto F. Progressive differentiation and selection of the fittest in the immune response. Nat Rev Immunol. 2002;2:982–987. - PubMed
-
- King CG, et al. TRAF6 is a T cell-intrinsic negative regulator required for the maintenance of immune homeostasis. Nat Med. 2006;12:1088–1092. - PubMed
-
- Bachmaier K, et al. Negative regulation of lymphocyte activation and autoimmunity by the molecular adaptor Cbl-b. Nature. 2000;403:211–216. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
