

Size and shape of protein molecules at the nanometer level determined by sedimentation, gel filtration, and electron microscopy
- PMID: 19495910
- PMCID: PMC3055910
- DOI: 10.1007/s12575-009-9008-x
Size and shape of protein molecules at the nanometer level determined by sedimentation, gel filtration, and electron microscopy
Abstract
An important part of characterizing any protein molecule is to determine its size and shape. Sedimentation and gel filtration are hydrodynamic techniques that can be used for this medium resolution structural analysis. This review collects a number of simple calculations that are useful for thinking about protein structure at the nanometer level. Readers are reminded that the Perrin equation is generally not a valid approach to determine the shape of proteins. Instead, a simple guideline is presented, based on the measured sedimentation coefficient and a calculated maximum S, to estimate if a protein is globular or elongated. It is recalled that a gel filtration column fractionates proteins on the basis of their Stokes radius, not molecular weight. The molecular weight can be determined by combining gradient sedimentation and gel filtration, techniques available in most biochemistry laboratories, as originally proposed by Siegel and Monte. Finally, rotary shadowing and negative stain electron microscopy are powerful techniques for resolving the size and shape of single protein molecules and complexes at the nanometer level. A combination of hydrodynamics and electron microscopy is especially powerful.
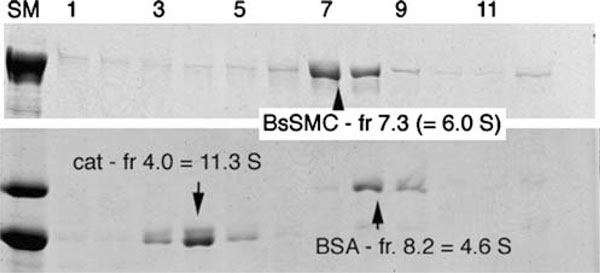
Figures





Similar articles
-
Determining the subunit structure of phosphodiesterases using gel filtration and sucrose density gradient centrifugation.Methods Mol Biol. 2005;307:167-80. doi: 10.1385/1-59259-839-0:167. Methods Mol Biol. 2005. PMID: 15988063
-
Structure and hydrodynamic properties of plectin molecules.J Mol Biol. 1987 Dec 5;198(3):515-31. doi: 10.1016/0022-2836(87)90297-x. J Mol Biol. 1987. PMID: 3430617
-
Size and shape of bovine interphotoreceptor retinoid-binding protein by electron microscopy and hydrodynamic analysis.J Biol Chem. 1987 Sep 25;262(27):13198-203. J Biol Chem. 1987. PMID: 3654607
-
Determination of Hydrodynamic Radius of Proteins by Size Exclusion Chromatography.Bio Protoc. 2017 Apr 20;7(8):e2230. doi: 10.21769/BioProtoc.2230. eCollection 2017 Apr 20. Bio Protoc. 2017. PMID: 34541230 Free PMC article.
-
Molecular weight standards for calibration of gel filtration and sodium dodecyl sulfate-polyacrylamide gel electrophoresis: ferritin and apoferritin.Anal Biochem. 1987 Nov 1;166(2):235-45. doi: 10.1016/0003-2697(87)90570-7. Anal Biochem. 1987. PMID: 3324819 Review.
Cited by
-
Structural Insights into N-terminal IgV Domain of BTNL2, a T Cell Inhibitory Molecule, Suggests a Non-canonical Binding Interface for Its Putative Receptors.J Mol Biol. 2020 Nov 6;432(22):5938-5950. doi: 10.1016/j.jmb.2020.09.013. Epub 2020 Oct 5. J Mol Biol. 2020. PMID: 32976909 Free PMC article.
-
Standardized bacteriophage purification for personalized phage therapy.Nat Protoc. 2020 Sep;15(9):2867-2890. doi: 10.1038/s41596-020-0346-0. Epub 2020 Jul 24. Nat Protoc. 2020. PMID: 32709990
-
Engineered nanodrug targeting oxidative stress for treatment of acute kidney injury.Exploration (Beijing). 2023 Jul 20;3(6):20220148. doi: 10.1002/EXP.20220148. eCollection 2023 Dec. Exploration (Beijing). 2023. PMID: 38264689 Free PMC article. Review.
-
Cellular imaging of endosome entrapped small gold nanoparticles.MethodsX. 2015 Jun 10;2:306-15. doi: 10.1016/j.mex.2015.06.001. eCollection 2015. MethodsX. 2015. PMID: 26151001 Free PMC article.
-
Dual-Angle Interferometric Scattering Microscopy for Optical Multiparametric Particle Characterization.Nano Lett. 2024 Feb 14;24(6):1874-1881. doi: 10.1021/acs.nanolett.3c03539. Epub 2024 Jan 31. Nano Lett. 2024. PMID: 38295760 Free PMC article.
References
-
- Sober HA. Handbook of Biochemistry. The Chemical Rubber Co., Cleveland, OH; 1966.
-
- Hesterberg LK, Lee JC, Erickson HP. Structural properties of an active form of rabbit muscle phosphofructokinase. J Biol Chem. 1981;256:9724–9730. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
