

Mice lacking Pctp /StarD2 exhibit increased adaptive thermogenesis and enlarged mitochondria in brown adipose tissue
- PMID: 19502644
- PMCID: PMC2759827
- DOI: 10.1194/jlr.M900013-JLR200
Mice lacking Pctp /StarD2 exhibit increased adaptive thermogenesis and enlarged mitochondria in brown adipose tissue
Abstract
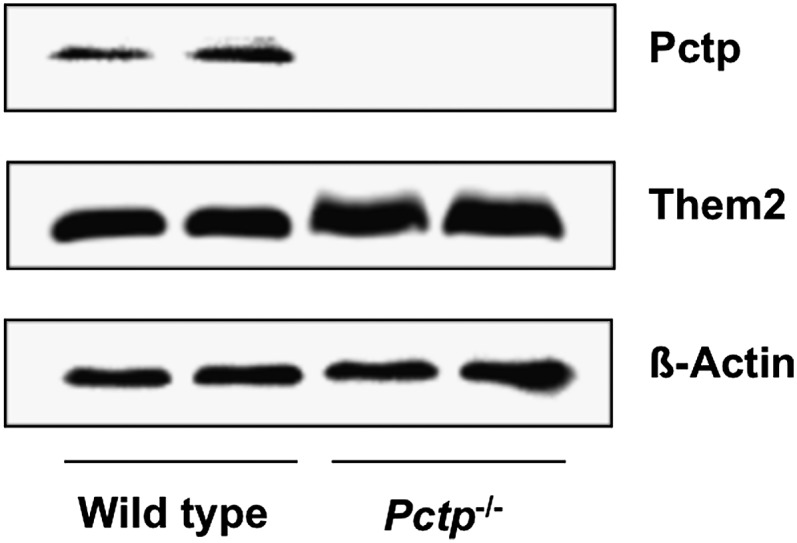
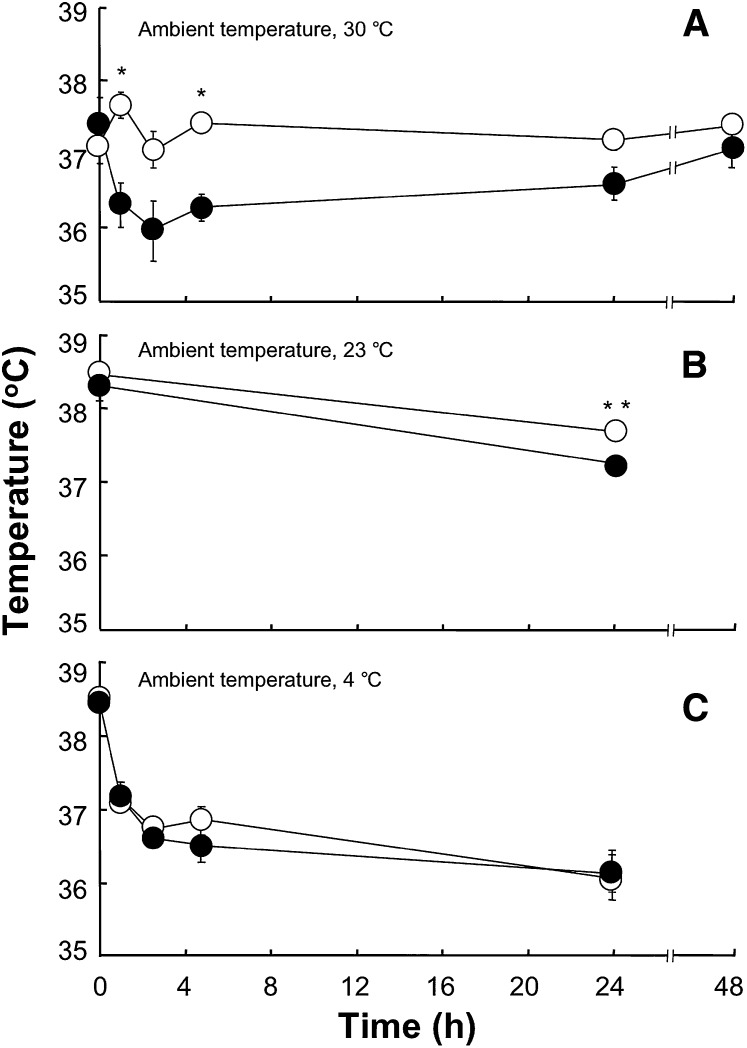
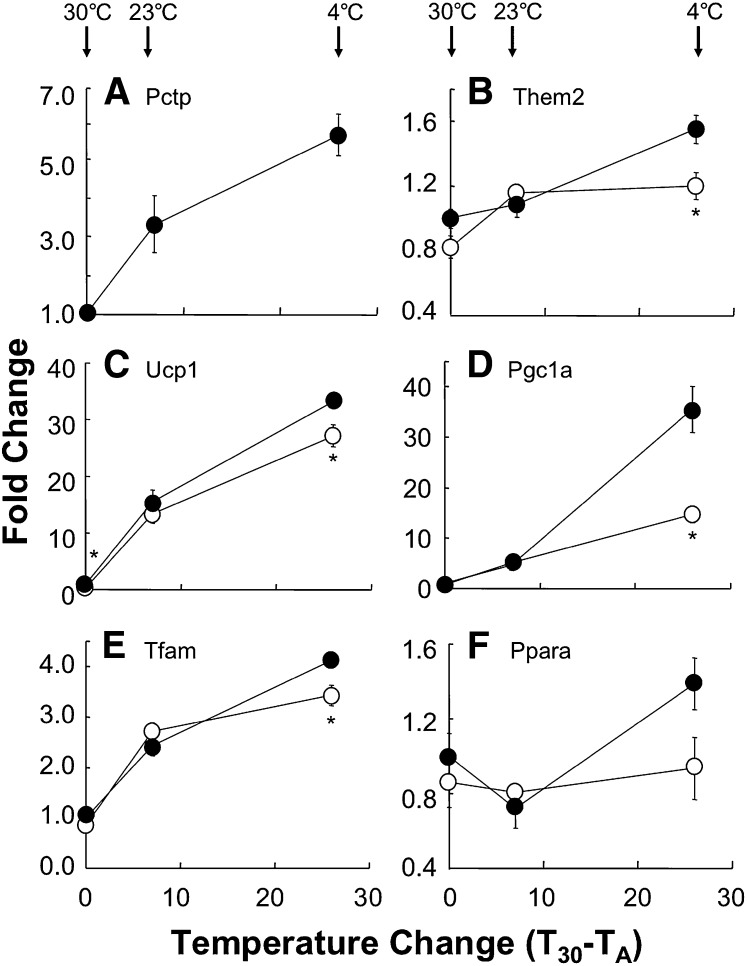
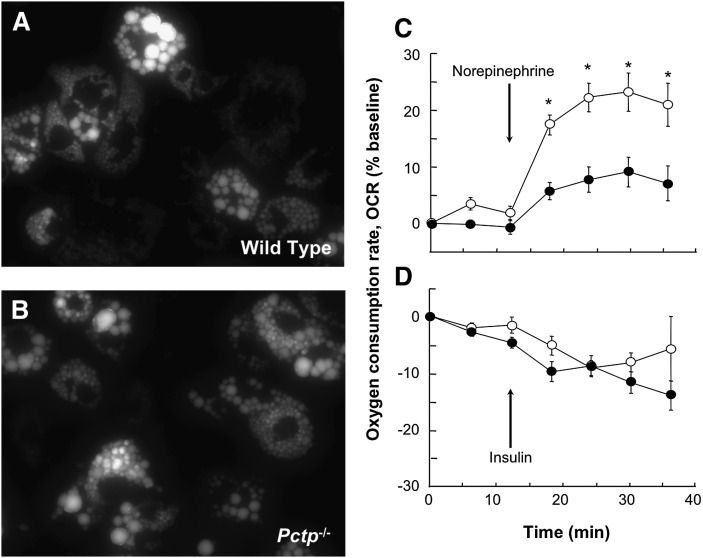
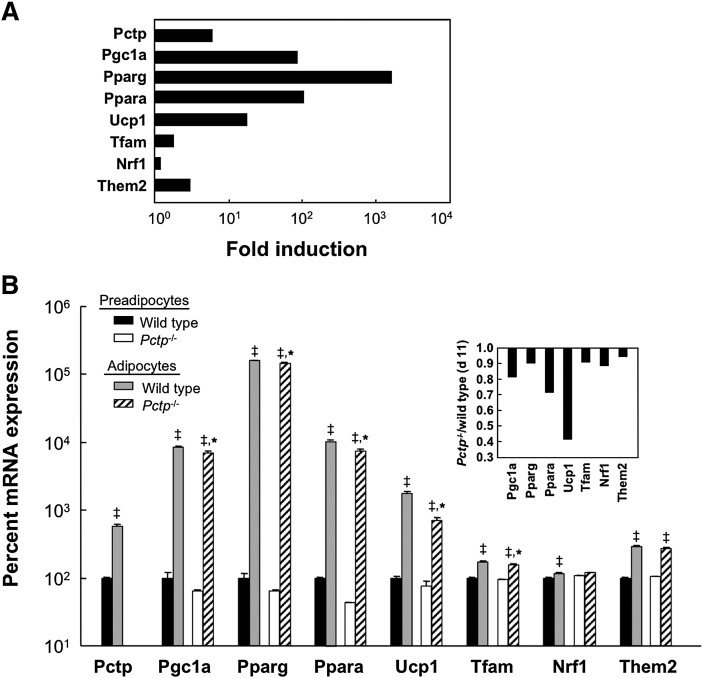
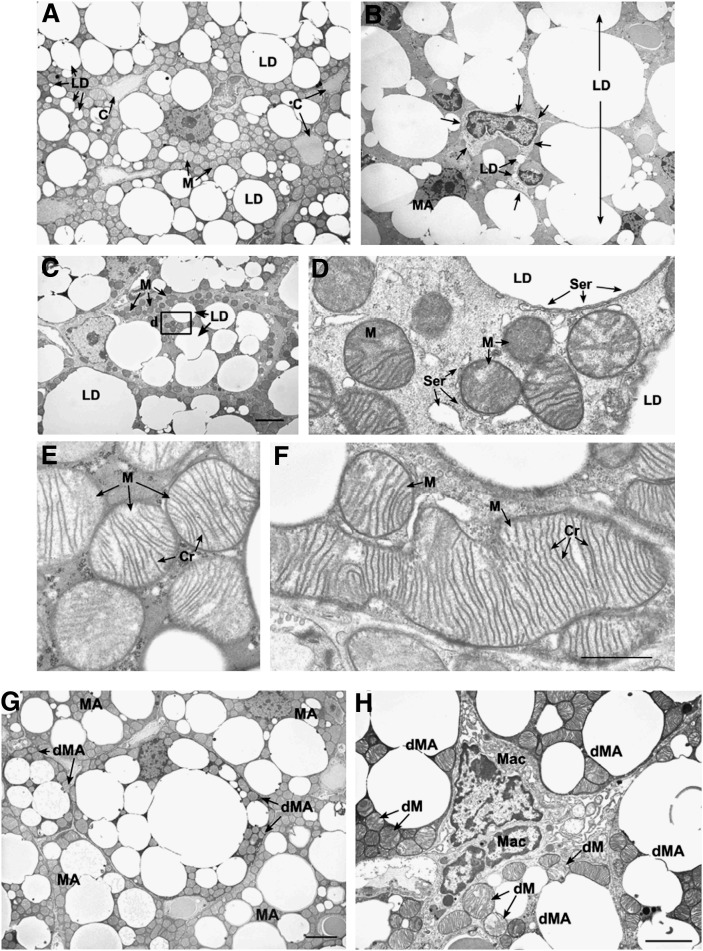
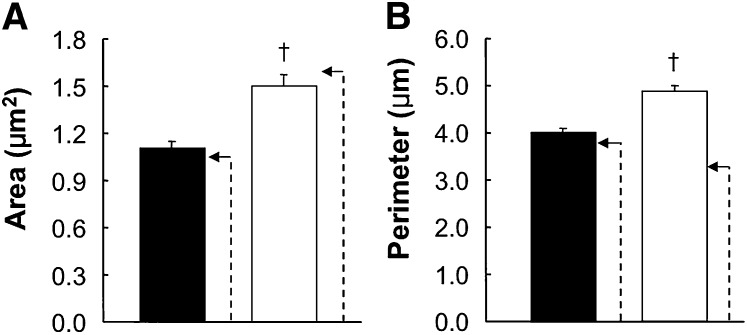
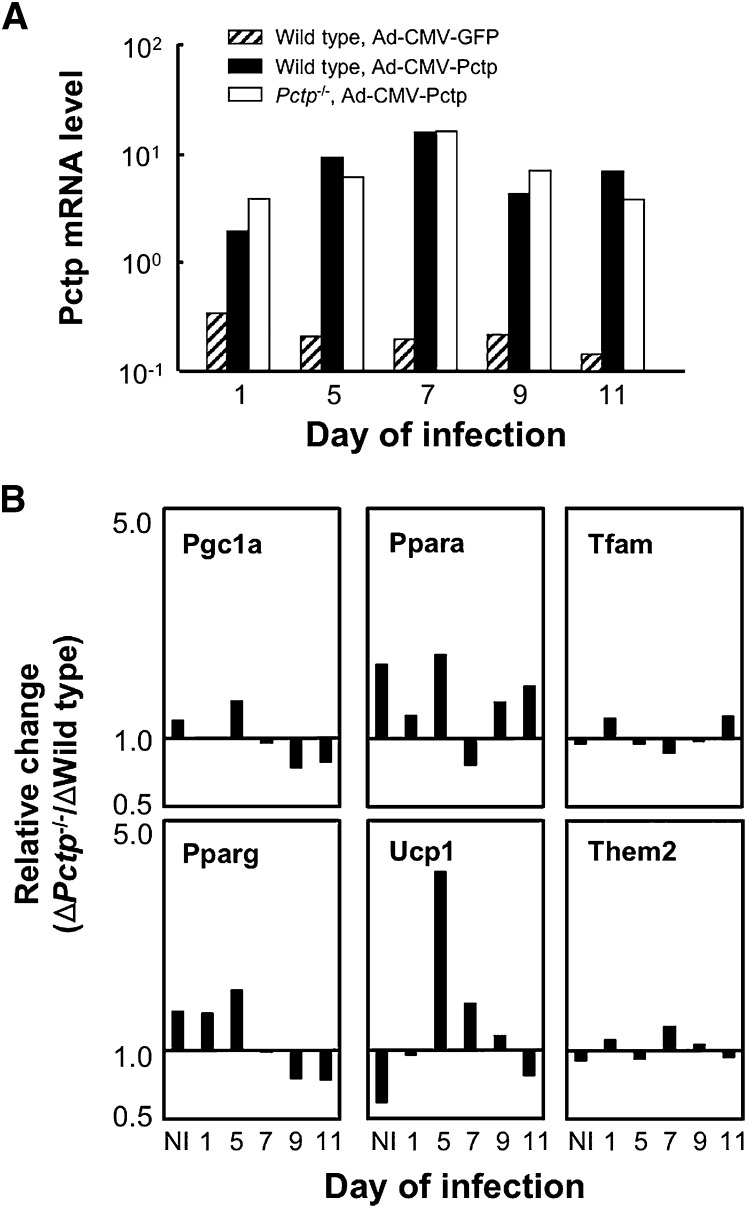
Pctp(-/-) mice that lack phosphatidylcholine transfer protein (Pctp) exhibit a marked shift toward utilization of fatty acids for oxidative phosphorylation, suggesting that Pctp may regulate the entry of fatty acyl-CoAs into mitochondria. Here, we examined the influence of Pctp expression on the function and structure of brown adipose tissue (BAT), a mitochondrial-rich, oxidative tissue that mediates nonshivering thermogenesis. Consistent with increased thermogenesis, Pctp(-/-) mice exhibited higher core body temperatures than wild-type controls at room temperature. During a 24 h cold challenge, Pctp(-/-) mice defended core body temperature efficiently enough that acute, full activation of BAT thermogenic genes did not occur. Brown adipocytes lacking Pctp harbored enlarged and elongated mitochondria. Consistent with increased fatty acid utilization, brown adipocytes cultured from Pctp(-/-) mice exhibited higher oxygen consumption rates in response to norepinephrine. The absence of Pctp expression during brown adipogenesis in vitro altered the expression of key transcription factors, which could be corrected by adenovirus-mediated overexpression of Pctp early but not late during the differentiation. Collectively, these findings support a key role for Pctp in limiting mitochondrial oxidation of fatty acids and thus regulating adaptive thermogenesis in BAT.
Figures
References
-
- Ponting C. P., Aravind L. 1999. START: a lipid-binding domain in StAR, HD-ZIP and signalling proteins. Trends Biochem. Sci. 24: 130–132. - PubMed
-
- Roderick S. L., Chan W. W., Agate D. S., Olsen L. R., Vetting M. W., Rajashankar K. R., Cohen D. E. 2002. Structure of human phosphatidylcholine transfer protein in complex with its ligand. Nat. Struct. Biol. 9: 507–511. - PubMed
-
- Alpy F., Tomasetto C. 2005. Give lipids a START: the StAR-related lipid transfer (START) domain in mammals. J. Cell Sci. 118: 2791–2801. - PubMed
-
- Soccio R. E., Breslow J. L. 2003. StAR-related lipid transfer (START) proteins: mediators of intracellular lipid metabolism. J. Biol. Chem. 278: 22183–22186. - PubMed
-
- Tsujishita Y., Hurley J. H. 2000. Structure and lipid transport mechanism of a StAR-related domain. Nat. Struct. Biol. 7: 408–414. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
