

ATXR5 and ATXR6 are H3K27 monomethyltransferases required for chromatin structure and gene silencing
- PMID: 19503079
- PMCID: PMC2754316
- DOI: 10.1038/nsmb.1611
ATXR5 and ATXR6 are H3K27 monomethyltransferases required for chromatin structure and gene silencing
Abstract
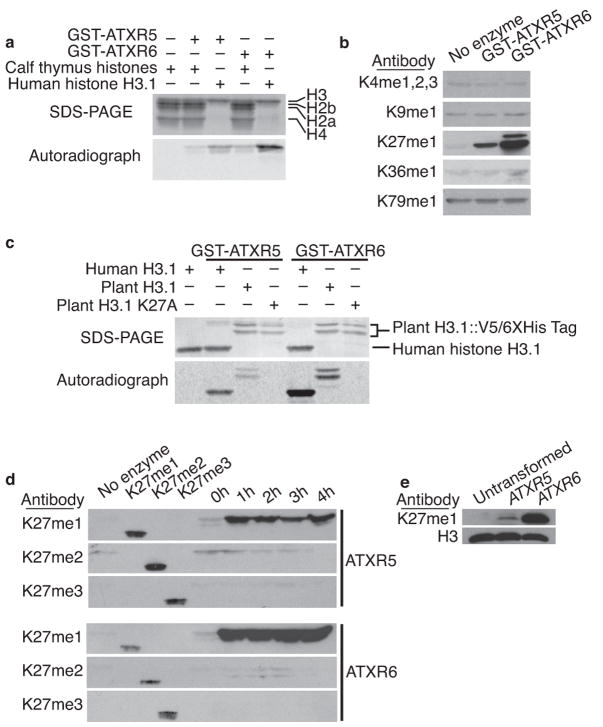
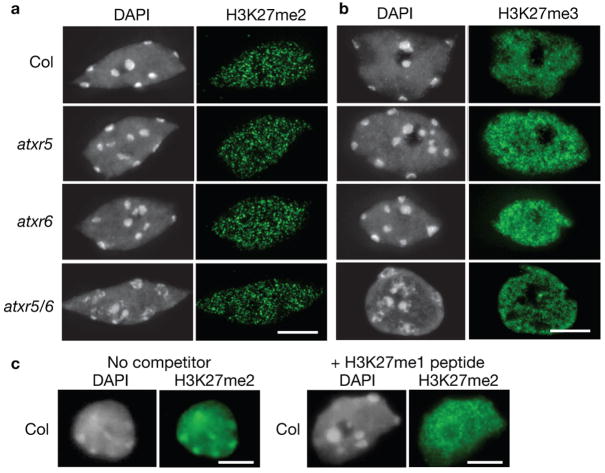
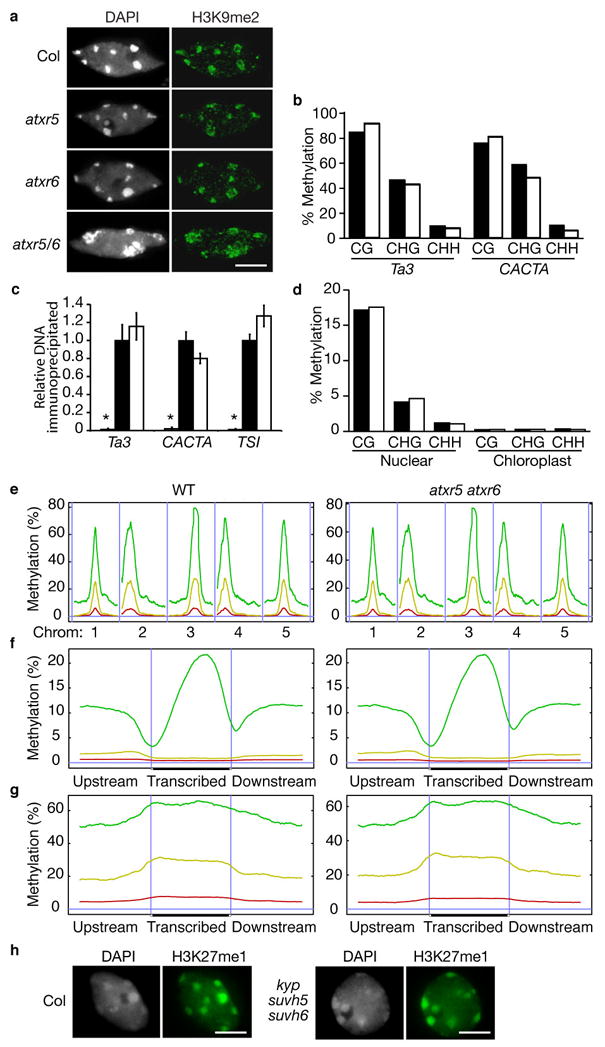
Constitutive heterochromatin in Arabidopsis thaliana is marked by repressive chromatin modifications, including DNA methylation, histone H3 dimethylation at Lys9 (H3K9me2) and monomethylation at Lys27 (H3K27me1). The enzymes catalyzing DNA methylation and H3K9me2 have been identified; alterations in these proteins lead to reactivation of silenced heterochromatic elements. The enzymes responsible for heterochromatic H3K27me1, in contrast, remain unknown. Here we show that the divergent SET-domain proteins ARABIDOPSIS TRITHORAX-RELATED PROTEIN 5 (ATXR5) and ATXR6 have H3K27 monomethyltransferase activity, and atxr5 atxr6 double mutants have reduced H3K27me1 in vivo and show partial heterochromatin decondensation. Mutations in atxr5 and atxr6 also lead to transcriptional activation of repressed heterochromatic elements. Notably, H3K9me2 and DNA methylation are unaffected in double mutants. These results indicate that ATXR5 and ATXR6 form a new class of H3K27 methyltransferases and that H3K27me1 represents a previously uncharacterized pathway required for transcriptional repression in Arabidopsis.
Conflict of interest statement
The authors declare that they have no competing financial interests.
Figures


References
-
- Li B, Carey M, Workman JL. The role of chromatin during transcription. Cell. 2007;128:707–19. - PubMed
-
- Maluszynska J, Heslop-Harrison JS. Localization of tandemly repeated DNA sequences in Arabidopsis thaliana. Plant J. 1991;1:159–166.
-
- Johnson L, Cao X, Jacobsen S. Interplay between two epigenetic marks. DNA methylation and histone H3 lysine 9 methylation. Curr Biol. 2002;12:1360–7. - PubMed
-
- Probst AV, Fransz PF, Paszkowski J, Mittelsten Scheid O. Two means of transcriptional reactivation within heterochromatin. Plant J. 2003;33:743–9. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
