

At the crossroads of homoeostasis and disease: roles of the PACS proteins in membrane traffic and apoptosis
- PMID: 19505291
- PMCID: PMC4303049
- DOI: 10.1042/BJ20081016
At the crossroads of homoeostasis and disease: roles of the PACS proteins in membrane traffic and apoptosis
Abstract
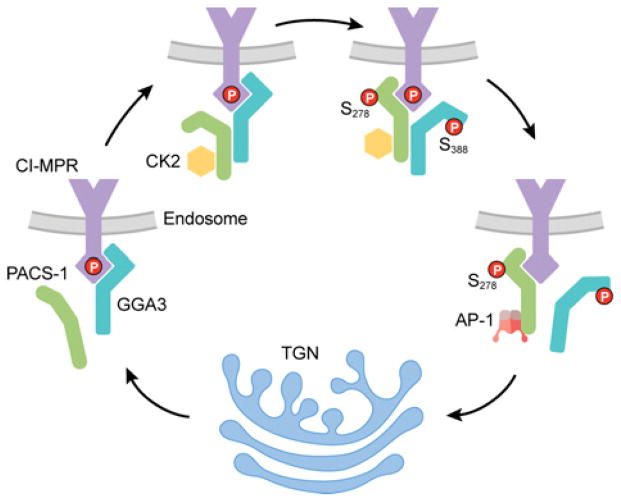
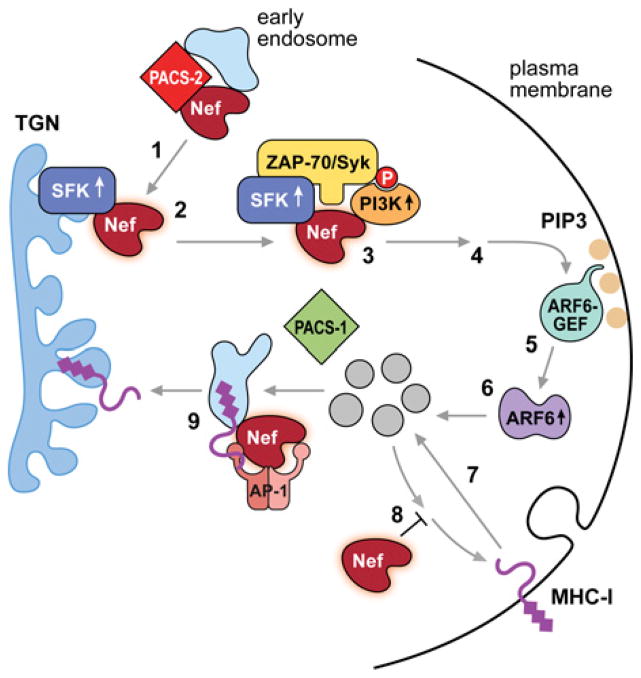
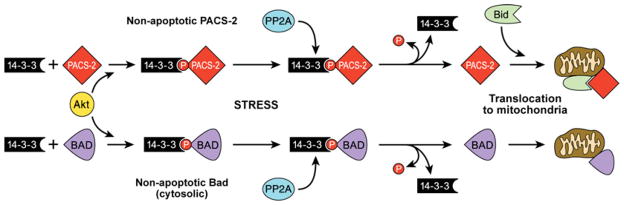
The endomembrane system in mammalian cells has evolved over the past two billion years from a simple endocytic pathway in a single-celled primordial ancestor to complex networks supporting multicellular structures that form metazoan tissue and organ systems. The increased organellar complexity of metazoan cells requires additional trafficking machinery absent in yeast or other unicellular organisms to maintain organ homoeostasis and to process the signals that control proliferation, differentiation or the execution of cell death programmes. The PACS (phosphofurin acidic cluster sorting) proteins are one such family of multifunctional membrane traffic regulators that mediate organ homoeostasis and have important roles in diverse pathologies and disease states. This review summarizes our current knowledge of the PACS proteins, including their structure and regulation in cargo binding, their genetics, their roles in secretory and endocytic pathway traffic, interorganellar communication and how cell-death signals reprogramme the PACS proteins to regulate apoptosis. We also summarize our current understanding of how PACS genes are dysregulated in cancer and how viral pathogens ranging from HIV-1 to herpesviruses have evolved to usurp the PACS sorting machinery to promote virus assembly, viral spread and immunoevasion.
Figures
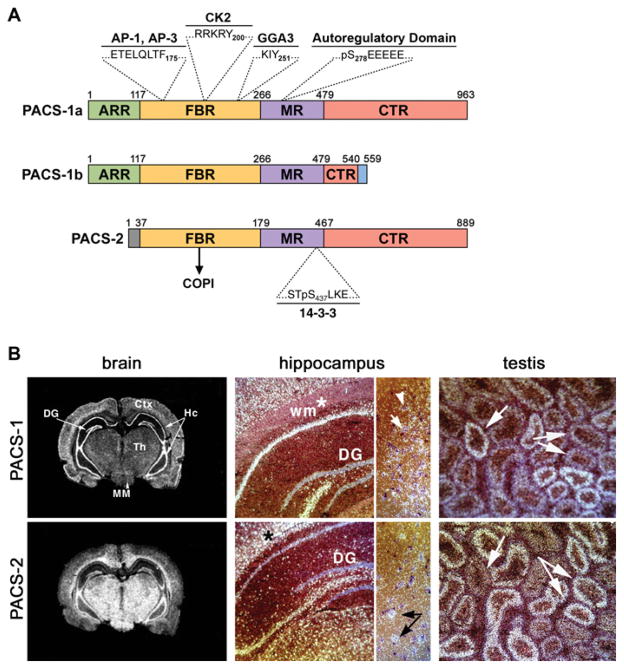
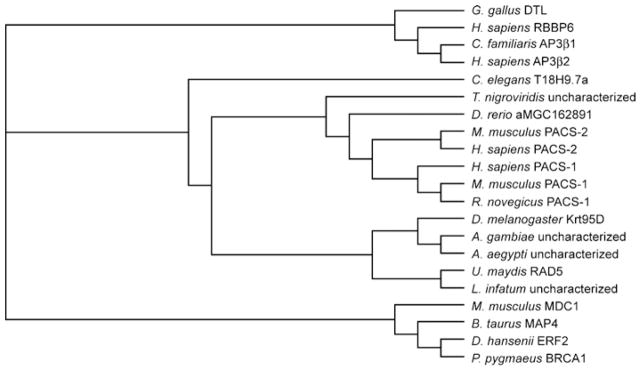
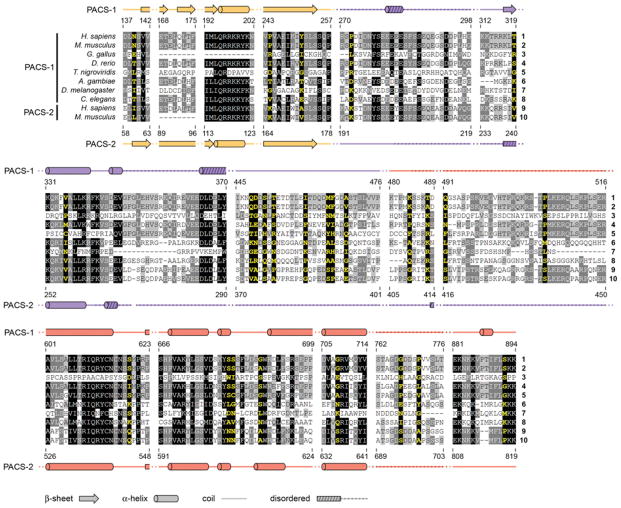
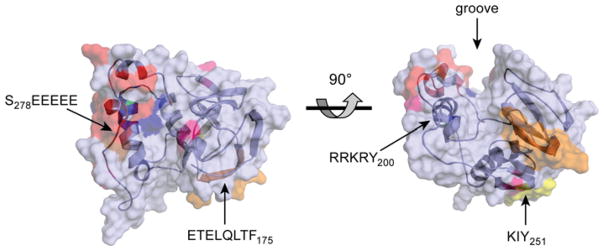
References
-
- Lee MC, Miller EA, Goldberg J, Orci L, Schekman R. Bi-directional protein transport between the ER and Golgi. Annu Rev Cell Dev Biol. 2004;20:87–123. - PubMed
-
- McMahon HT, Mills IG. COP and clathrin-coated vesicle budding: different pathways, common approaches. Curr Opin Cell Biol. 2004;16:379–391. - PubMed
-
- Palade G. Intracellular aspects of the process of protein synthesis. Science. 1975;189:347–358. - PubMed
-
- Fewell SW, Travers KJ, Weissman JS, Brodsky JL. The action of molecular chaperones in the early secretory pathway. Annu Rev Genet. 2001;35:149–191. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous