

Evolution and diversity of glutaredoxins in photosynthetic organisms
- PMID: 19506802
- PMCID: PMC11115520
- DOI: 10.1007/s00018-009-0054-y
Evolution and diversity of glutaredoxins in photosynthetic organisms
Abstract
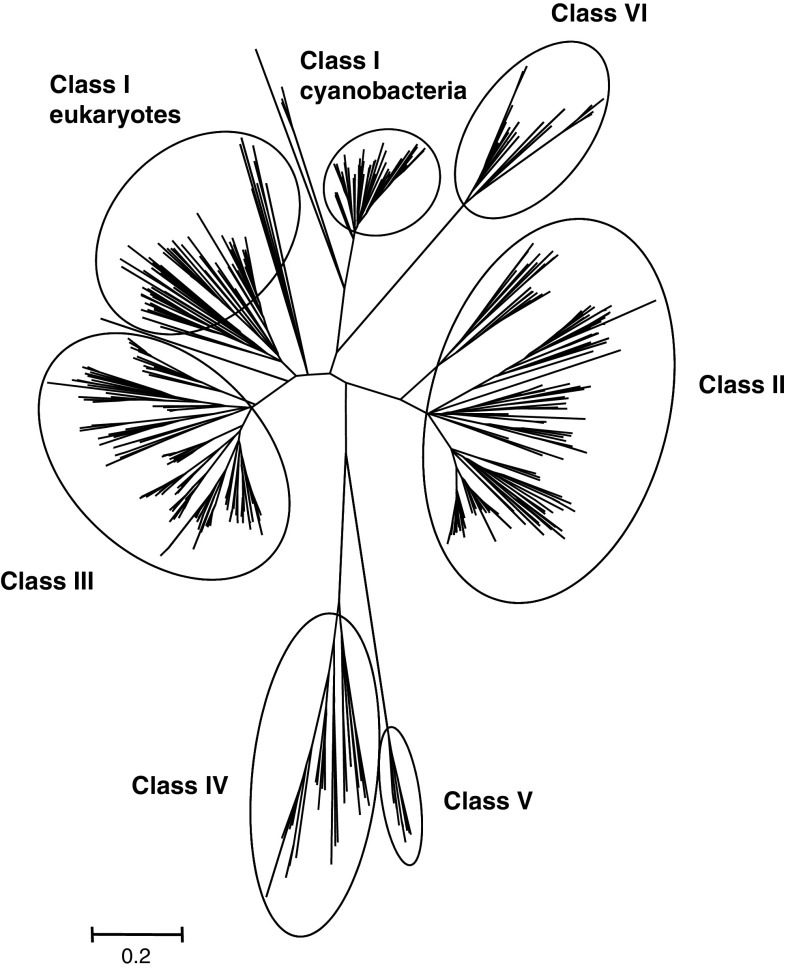
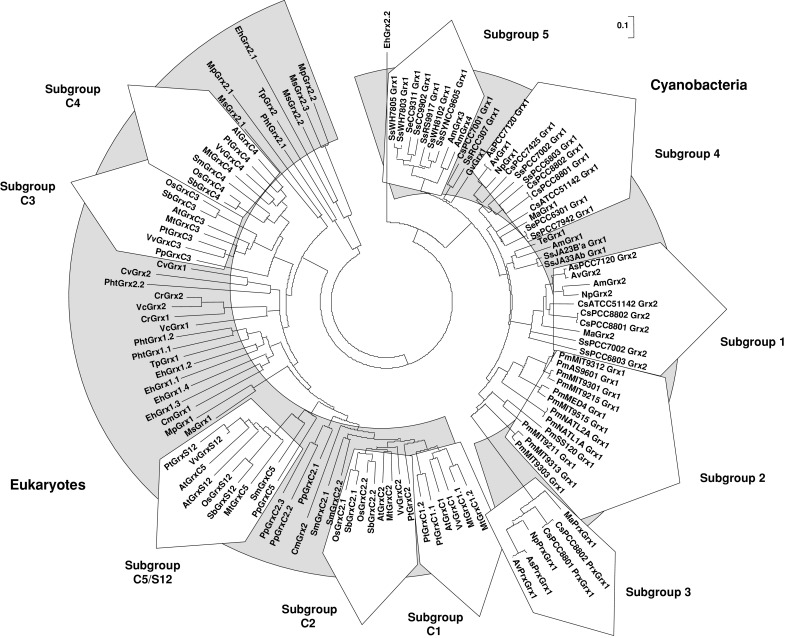
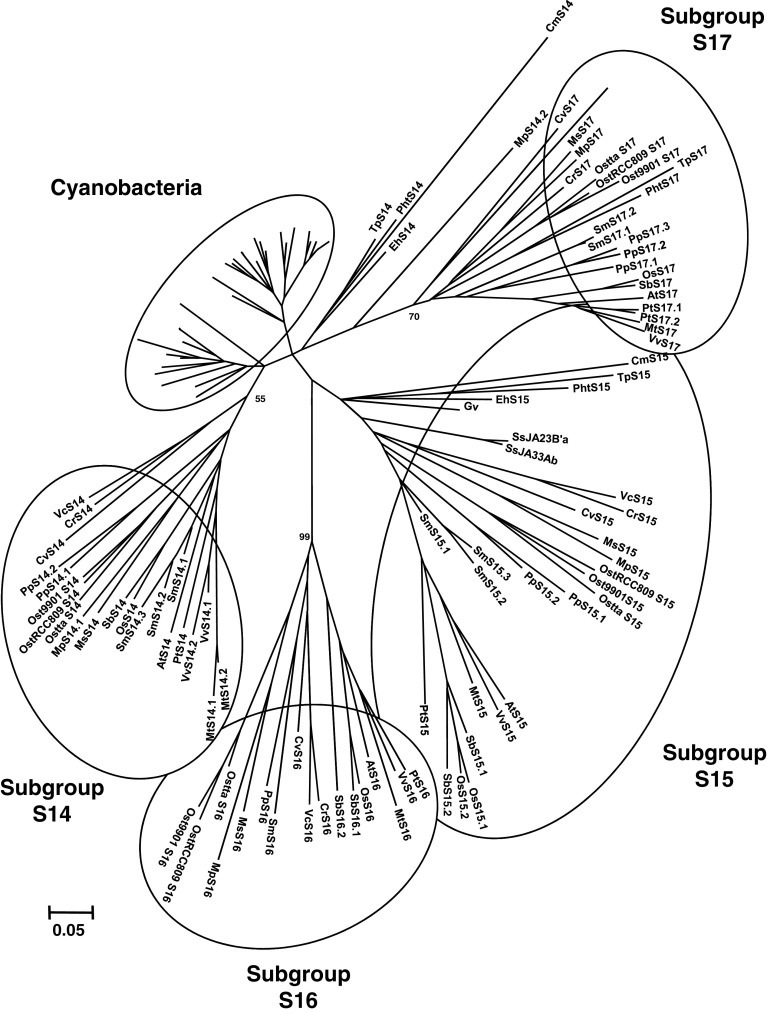
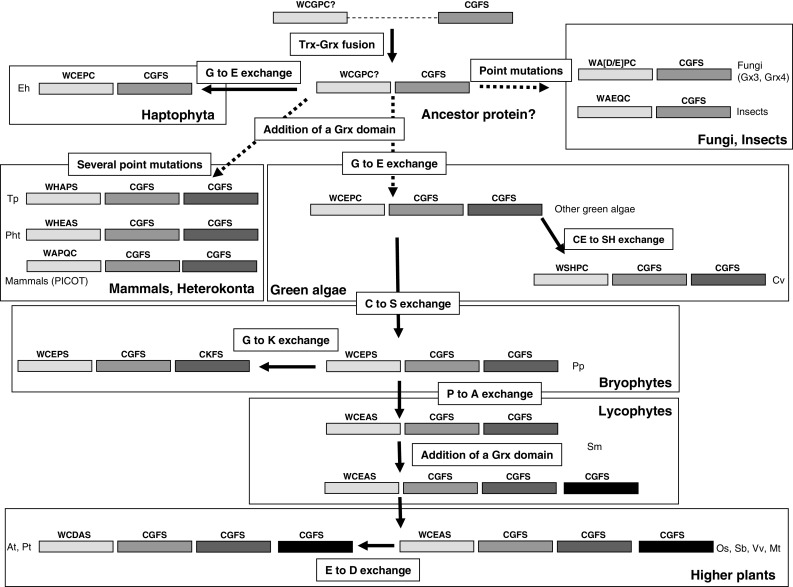
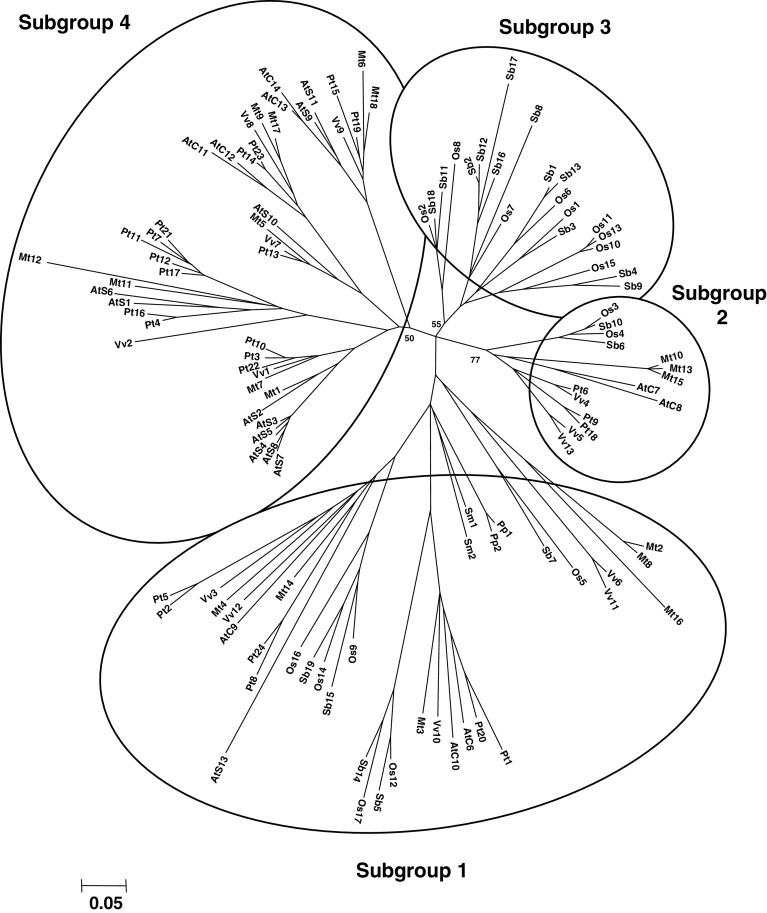
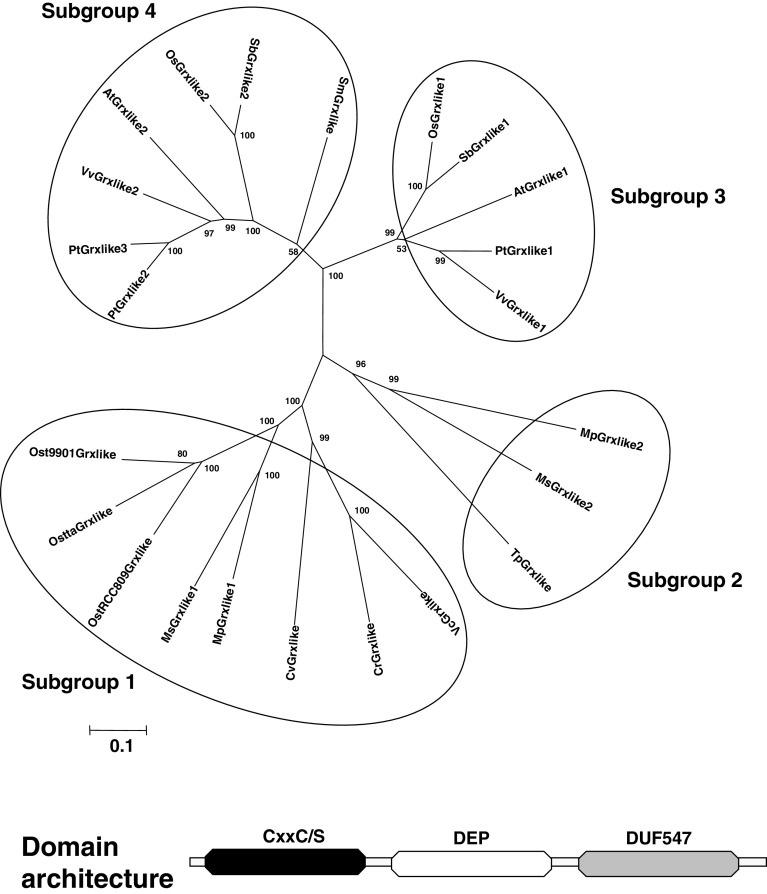
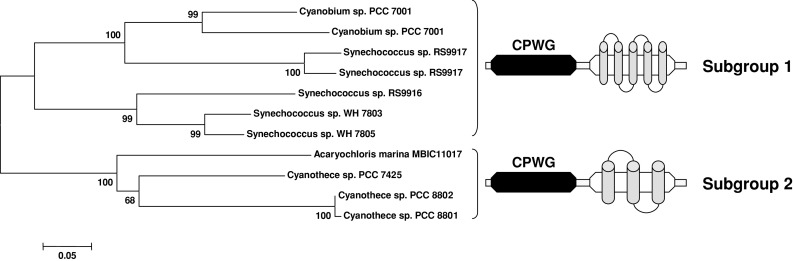
The genome sequencing of prokaryotic and eukaryotic photosynthetic organisms enables a comparative genomic study of the glutaredoxin (Grx) family. The analysis of 58 genomes, using a specific motif composed of the active site sequence and of amino acids involved in glutathione binding, led to an updated classification of Grxs into six classes. Only two classes (I and II) are common to all photosynthetic organisms. Eukaryotes and cyanobacteria have two specific Grx classes (classes III and IV and classes V and VI, respectively). The classes IV, V and VI have not yet been identified and contain multimodular Grx fusions. In addition, putative Grx partners were identified from the presence of fusion proteins, the conservation of gene order in bacterial operons, and the gene co-occurrence. The genes encoding class II Grxs and BolA/YrbA proteins are frequently adjacent, in the same transcriptional orientation in prokaryote genomes and present in the same organisms.
Figures


References
-
- Fernandes AP, Fladvad M, Berndt C, Andresen C, Lillig CH, Neubauer P, Sunnerhagen M, Holmgren A, Vlamis-Gardikas A. A novel monothiol glutaredoxin (Grx4) from Escherichia coli can serve as a substrate for thioredoxin reductase. J Biol Chem. 2005;280:24544–24552. doi: 10.1074/jbc.M500678200. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
