

Genome-wide analysis of mono-, di- and trimethylation of histone H3 lysine 4 in Arabidopsis thaliana
- PMID: 19508735
- PMCID: PMC2718496
- DOI: 10.1186/gb-2009-10-6-r62
Genome-wide analysis of mono-, di- and trimethylation of histone H3 lysine 4 in Arabidopsis thaliana
Abstract
Background: Post-translational modifications of histones play important roles in maintaining normal transcription patterns by directly or indirectly affecting the structural properties of the chromatin. In plants, methylation of histone H3 lysine 4 (H3K4me) is associated with genes and required for normal plant development.
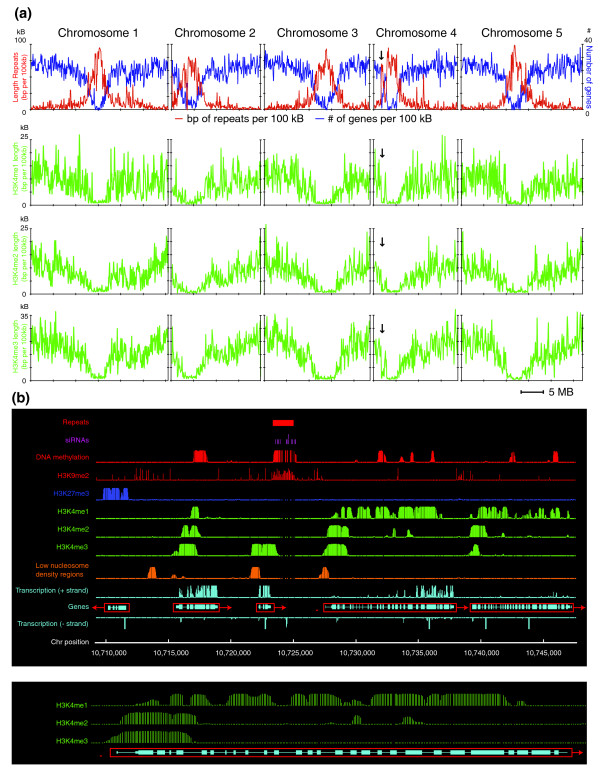
Results: We have characterized the genome-wide distribution patterns of mono-, di- and trimethylation of H3K4 (H3K4me1, H3K4me2 and H3K4me3, respectively) in Arabidopsis thaliana seedlings using chromatin immunoprecipitation and high-resolution whole-genome tiling microarrays (ChIP-chip). All three types of H3K4me are found to be almost exclusively genic, and two-thirds of Arabidopsis genes contain at least one type of H3K4me. H3K4me2 and H3K4me3 accumulate predominantly in promoters and 5' genic regions, whereas H3K4me1 is distributed within transcribed regions. In addition, H3K4me3-containing genes are highly expressed with low levels of tissue specificity, but H3K4me1 or H3K4me2 may not be directly involved in transcriptional activation. Furthermore, the preferential co-localization of H3K4me3 and H3K27me3 found in mammals does not appear to occur in plants at a genome-wide level, but H3K4me2 and H3K27me3 co-localize at a higher-than-expected frequency. Finally, we found that H3K4me2/3 and DNA methylation appear to be mutually exclusive, but surprisingly, H3K4me1 is highly correlated with CG DNA methylation in the transcribed regions of genes.
Conclusions: H3K4me plays widespread roles in regulating gene expression in plants. Although many aspects of the mechanisms and functions of H3K4me appear to be conserved among all three kingdoms, we observed significant differences in the relationship between H3K4me and transcription or other epigenetic pathways in plants and mammals.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
