

Calcium elevation in mitochondria is the main Ca2+ requirement for mitochondrial permeability transition pore (mPTP) opening
- PMID: 19515844
- PMCID: PMC2742844
- DOI: 10.1074/jbc.M109.025353
Calcium elevation in mitochondria is the main Ca2+ requirement for mitochondrial permeability transition pore (mPTP) opening
Abstract
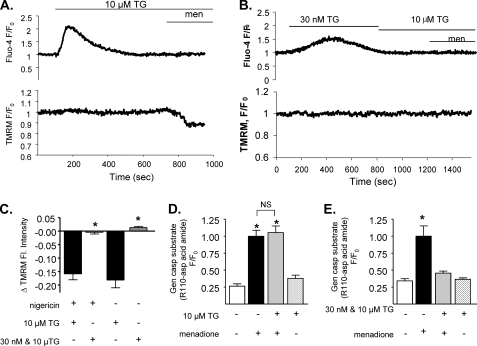
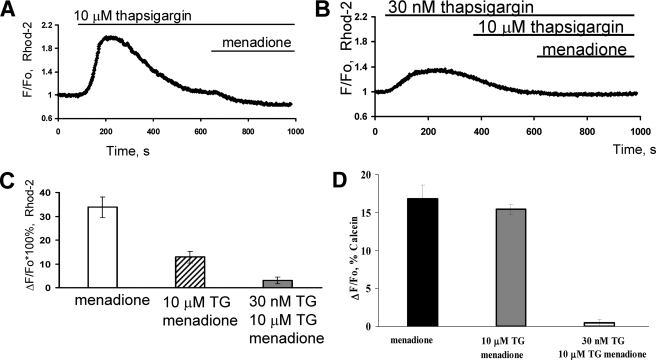
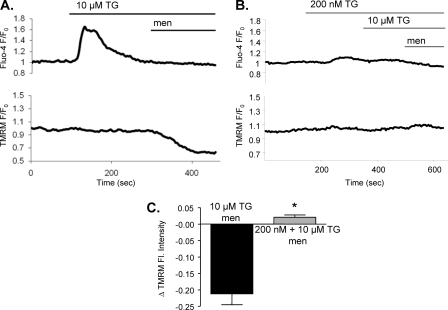
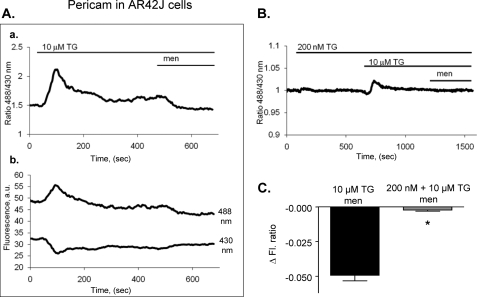
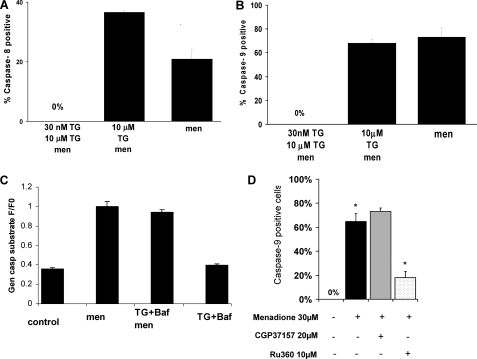
We have investigated in detail the role of intra-organelle Ca2+ content during induction of apoptosis by the oxidant menadione while changing and monitoring the Ca2+ load of endoplasmic reticulum (ER), mitochondria, and acidic organelles. Menadione causes production of reactive oxygen species, induction of oxidative stress, and subsequently apoptosis. In both pancreatic acinar and pancreatic tumor AR42J cells, menadione was found to induce repetitive cytosolic Ca2+ responses because of the release of Ca2+ from both ER and acidic stores. Ca2+ responses to menadione were accompanied by elevation of Ca2+ in mitochondria, mitochondrial depolarization, and mitochondrial permeability transition pore (mPTP) opening. Emptying of both the ER and acidic Ca2+ stores did not necessarily prevent menadione-induced apoptosis. High mitochondrial Ca2+ at the time of menadione application was the major factor determining cell fate. However, if mitochondria were prevented from loading with Ca2+ with 10 mum RU360, then caspase-9 activation did not occur irrespective of the content of other Ca2+ stores. These results were confirmed by ratiometric measurements of intramitochondrial Ca2+ with pericam. We conclude that elevated Ca2+ in mitochondria is the crucial factor in determining whether cells undergo oxidative stress-induced apoptosis.
Figures
References
-
- Thornberry N. A., Lazebnik Y. (1998) Science 281, 1312–1316 - PubMed
-
- Berridge M. J., Lipp P., Bootman M. D. (2000) Nat. Rev. Mol. Cell Biol. 1, 11–21 - PubMed
-
- Huang Y., Fischer J. E., Balasubramaniam A. (1996) Peptides 17, 497–502 - PubMed
-
- Gerasimenko J. V., Gerasimenko O. V., Palejwala A., Tepikin A. V., Petersen O. H., Watson A. J. M. (2002) J. Cell Science 115, 485–497 - PubMed
-
- Baumgartner H. K., Gerasimenko J. V., Thorne C., Ashurst L. H., Barrow S. L., Chvanov M. A., Gillies S., Criddle D. N., Tepikin A. V., Petersen O. H., Sutton R., Watson A. J., Gerasimenko O. V. (2007) Am. J. Physiol. Gastrointest. Liver Physiol. 293, G296–G307 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
