

Exercise induces cortical plasticity after neonatal spinal cord injury in the rat
- PMID: 19515923
- PMCID: PMC2743445
- DOI: 10.1523/JNEUROSCI.2474-08.2009
Exercise induces cortical plasticity after neonatal spinal cord injury in the rat
Abstract
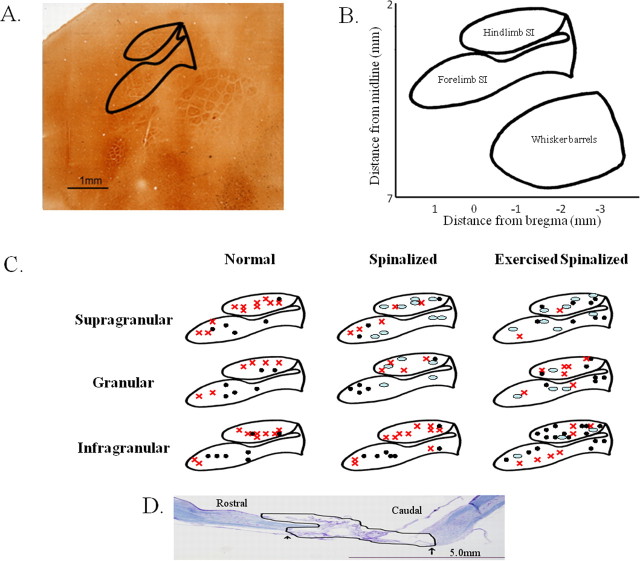
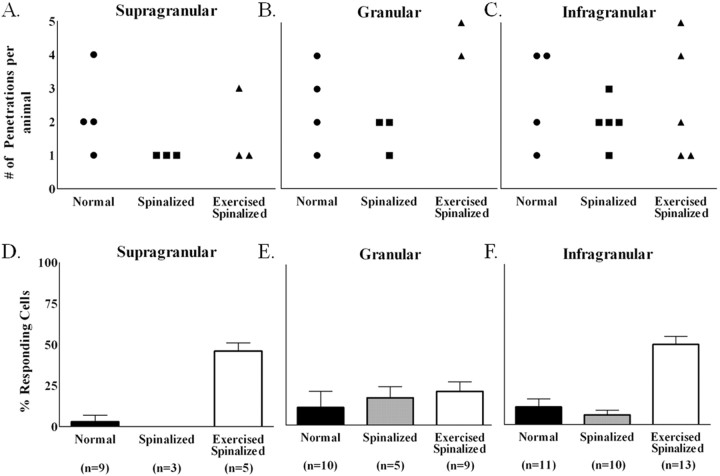
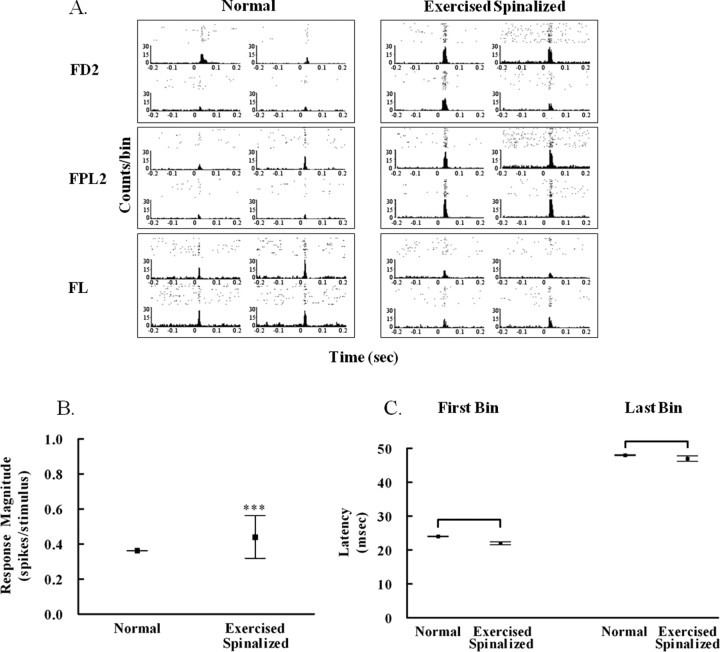
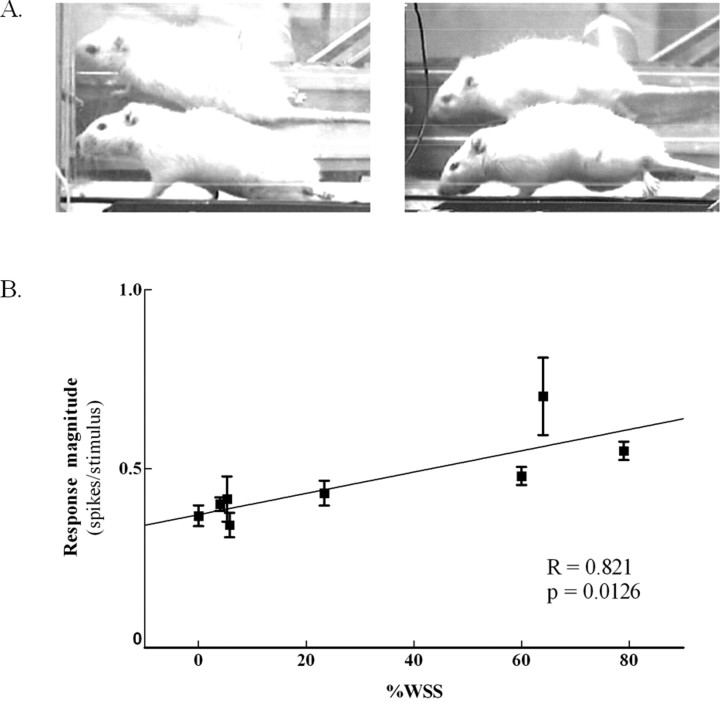
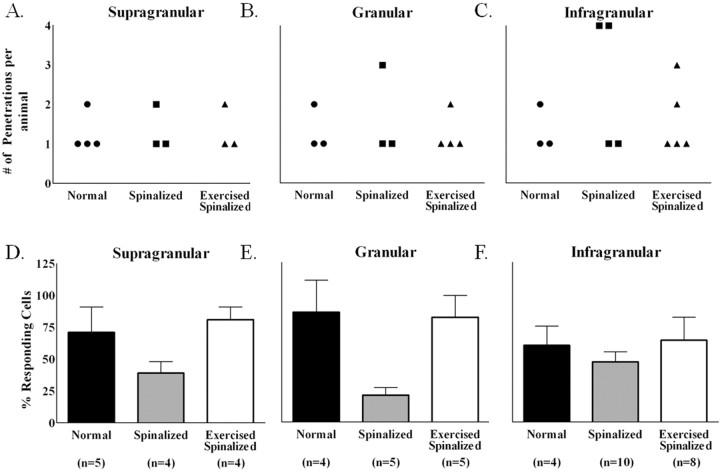
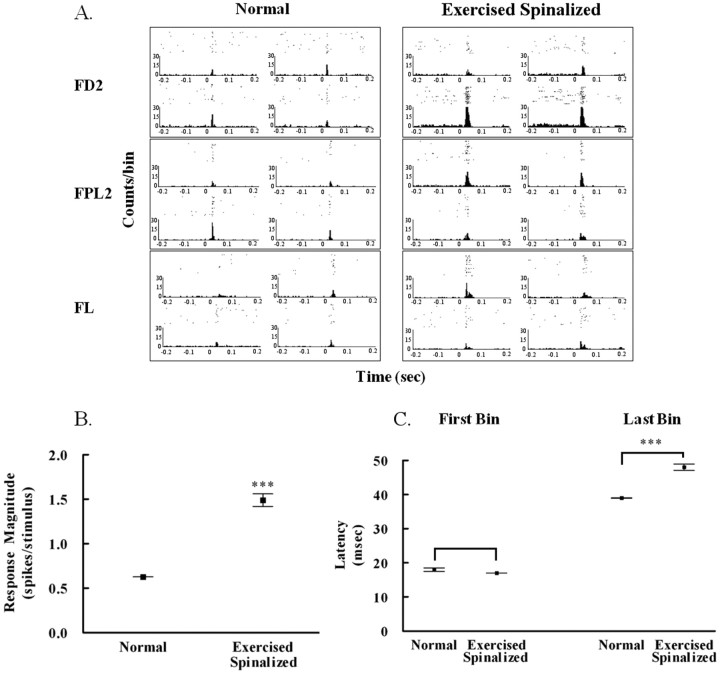
Exercise-induced cortical plasticity is associated with improved functional outcome after brain or nerve injury. Exercise also improves functional outcomes after spinal cord injury, but its effects on cortical plasticity are not known. The goal of this investigation was to study the effect of moderate exercise (treadmill locomotion, 3 min/d, 5 d/week) on the somatotopic organization of forelimb and hindlimb somatosensory cortex (SI) after neonatal thoracic transection. We used adult rats spinalized as neonates because some of these animals develop weight-supported stepping, and, therefore, the relationship between cortical plasticity and stepping could also be examined. Acute, single-neuron mapping was used to determine the percentage of cortical cells responding to cutaneous forelimb stimulation in normal, spinalized, and exercised spinalized rats. Multiple single-neuron recording from arrays of chronically implanted microwires examined the magnitude of response of these cells in normal and exercised spinalized rats. Our results show that exercise not only increased the percentage of responding cells in the hindlimb SI but also increased the magnitude of the response of these cells. This increase in response magnitude was correlated with behavioral outcome measures. In the forelimb SI, neonatal transection reduced the percentage of responding cells to forelimb stimulation, but exercise reversed this loss. This restoration in the percentage of responding cells after exercise was accompanied by an increase in their response magnitude. Therefore, the increase in responsiveness of hindlimb SI to forelimb stimulation after neonatal transection and exercise may be due, in part, to the effect of exercise on the forelimb SI.
Figures
Similar articles
-
Functional role of exercise-induced cortical organization of sensorimotor cortex after spinal transection.J Neurophysiol. 2011 Nov;106(5):2662-74. doi: 10.1152/jn.01017.2010. Epub 2011 Aug 24. J Neurophysiol. 2011. PMID: 21865438 Free PMC article.
-
Passive exercise of the hind limbs after complete thoracic transection of the spinal cord promotes cortical reorganization.PLoS One. 2013;8(1):e54350. doi: 10.1371/journal.pone.0054350. Epub 2013 Jan 22. PLoS One. 2013. PMID: 23349859 Free PMC article.
-
Role of cortical reorganization on the effect of 5-HT pharmacotherapy for spinal cord injury.Exp Neurol. 2013 Feb;240:17-27. doi: 10.1016/j.expneurol.2012.10.019. Epub 2012 Nov 13. Exp Neurol. 2013. PMID: 23159333
-
Transplants and neurotrophic factors increase regeneration and recovery of function after spinal cord injury.Prog Brain Res. 2002;137:257-73. doi: 10.1016/s0079-6123(02)37020-1. Prog Brain Res. 2002. PMID: 12440372 Review.
-
Cortical and subcortical plasticity in the brains of humans, primates, and rats after damage to sensory afferents in the dorsal columns of the spinal cord.Exp Neurol. 2008 Feb;209(2):407-16. doi: 10.1016/j.expneurol.2007.06.014. Epub 2007 Jul 6. Exp Neurol. 2008. PMID: 17692844 Free PMC article. Review.
Cited by
-
Trunk robot rehabilitation training with active stepping reorganizes and enriches trunk motor cortex representations in spinal transected rats.J Neurosci. 2015 May 6;35(18):7174-89. doi: 10.1523/JNEUROSCI.4366-14.2015. J Neurosci. 2015. PMID: 25948267 Free PMC article.
-
Cortical reorganization after spinal cord injury: always for good?Neuroscience. 2014 Dec 26;283:78-94. doi: 10.1016/j.neuroscience.2014.06.056. Epub 2014 Jul 2. Neuroscience. 2014. PMID: 24997269 Free PMC article. Review.
-
Functional recovery of stepping in rats after a complete neonatal spinal cord transection is not due to regrowth across the lesion site.Neuroscience. 2010 Mar 10;166(1):23-33. doi: 10.1016/j.neuroscience.2009.12.010. Epub 2009 Dec 17. Neuroscience. 2010. PMID: 20006680 Free PMC article.
-
Plasticity and alterations of trunk motor cortex following spinal cord injury and non-stepping robot and treadmill training.Exp Neurol. 2014 Jun;256:57-69. doi: 10.1016/j.expneurol.2014.03.012. Epub 2014 Apr 3. Exp Neurol. 2014. PMID: 24704619 Free PMC article.
-
Hindlimb Somatosensory Information Influences Trunk Sensory and Motor Cortices to Support Trunk Stabilization.Cereb Cortex. 2021 Oct 1;31(11):5165-5187. doi: 10.1093/cercor/bhab150. Cereb Cortex. 2021. PMID: 34165153 Free PMC article.
References
-
- Abraham WC, Bear MF. Metaplasticity: the plasticity of synaptic plasticity. Trends Neurosci. 1996;19:126–130. - PubMed
-
- Chapin JK. Laminar differences in sizes, shapes, and response profiles of cutaneous receptive fields in the rat SI cortex. Exp Brain Res. 1986;62:549–559. - PubMed
-
- Chapin JK, Sadeq M, Guise JL. Corticocortical connections within the primary somatosensory cortex of the rat. J Comp Neurol. 1987;263:326–346. - PubMed
-
- Chau CW, McKinley PA. Chronological observations of primary somatosensory cortical maps in kittens following low thoracic (T12) spinal cord transection at 2 weeks of age. Somatosens Mot Res. 1991;8:355–376. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical