

Arabidopsis potential calcium sensors regulate nitric oxide levels and the transition to flowering
- PMID: 19517005
- PMCID: PMC2634334
- DOI: 10.4161/psb.2.6.4695
Arabidopsis potential calcium sensors regulate nitric oxide levels and the transition to flowering
Abstract
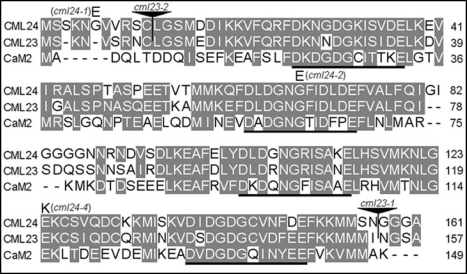
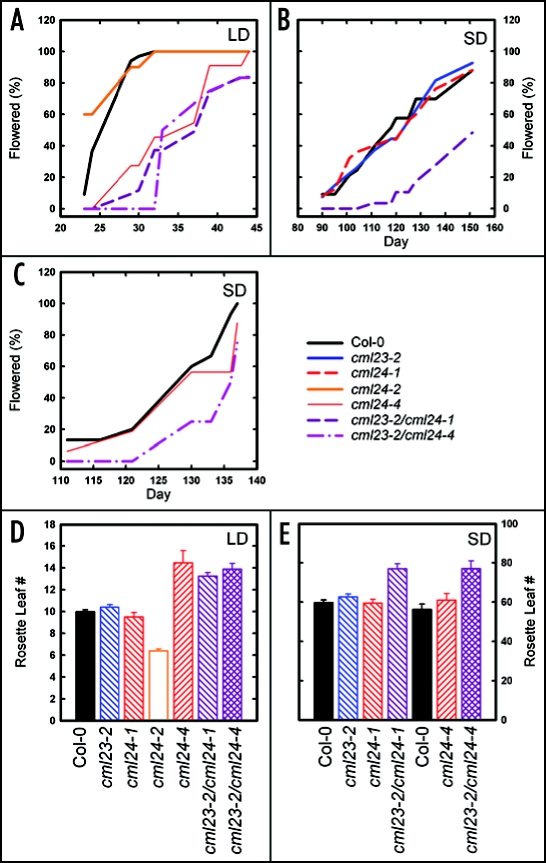
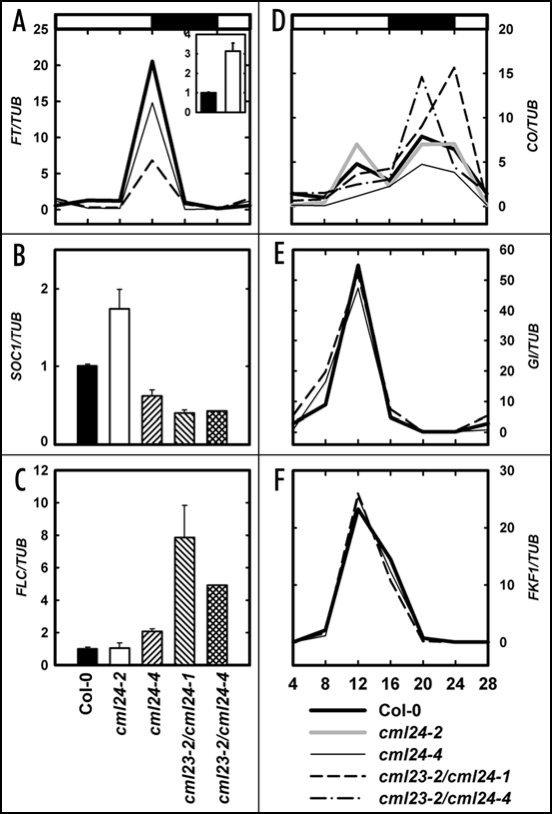
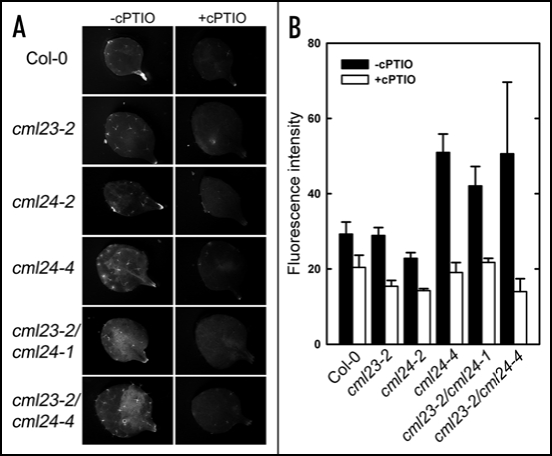
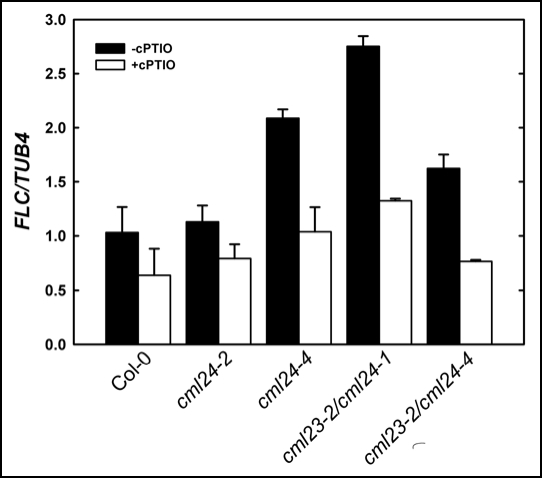
In plants, flowering is a critical developmental transition orchestrated by four regulatory pathways. Distinct alleles encoding mutant forms of the Arabidopsis potential calcium sensor CML24 cause alterations in flowering time. CML24 can act as a switch in the response to day length perception; loss-of-function cml24 mutants are late flowering under long days, whereas apparent gain of CML24 function results in early flowering. CML24 function is required for proper CONSTANS (CO) expression; components upstream of CO in the photoperiod pathway are largely unaffected in the cml24 mutants. In conjunction with CML23, a related calmodulin-like protein, CML24 also inhibits FLOWERING LOCUS C (FLC) expression and therefore impacts the autonomous regulatory pathway of the transition to flowering. Nitric oxide (NO) levels are elevated in cml23/cml24 double mutants and are largely responsible for FLC transcript accumulation. Therefore, CML23 and CML24 are potential calcium sensors that have partially overlapping function that may act to transduce calcium signals to regulate NO accumulation. In turn, NO levels influence the transition to flowering through both the photoperiod and autonomous regulatory pathways.
Keywords: calcium; calmodulin; cell signaling; flowering; nitric oxide.
Figures
References
-
- Suarez-Lopez P, Wheatley K, Robson F, Onouchi H, Valverde F, Coupland G. Constans mediates between the circadian clock and the control of flowering in Arabidopsis. Nature. 2001;410:1116–1120. - PubMed
-
- Yanovsky MJ, Kay SA. Molecular basis of seasonal time measurement in Arabidopsis. Nature. 2002;419:308–312. - PubMed
-
- Valverde F, Mouradov A, Soppe W, Ravenscroft D, Samach A, Coupland G. Photoreceptor regulation of CONSTANS protein in photoperiodic flowering. Science. 2004;303:1003–1006. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases